Fakulta sportovních studií Masarykovy univerzity
Doc. MUDr. Jitka Hanzlová, CSc., MUDr. Jan Hemza
Základy anatomie I
Základy anatomie II
Základy anatomie III
Základy anatomie IV – zobrazeno
Centrální nervový systém
Základní pojmy
Nervový systém je nejvyšší specializovaná a integrační soustava organismu živočichů.
Díky kvalitativní i kvantitativním podnětům působícím zvenku se postupně ve fylogenetickém i ontogenetickém vývoji vydiferencovala ze zevního listu – ektodermu nervová tkáň.
Má tři charakteristické vlastnosti:
- dráždivost, iritabilita – to je schopnost na základě energetických objemů reagovat a tyto energie transformovat.
- schopnost vodivosti - to je schopnost přenosu energetických objemů
- schopnost tvořit mezibuněčné vazby – interneronová synaptogeneze, a tím tvořit postupně různě hierarchizované sítě.
Složení nervové tkáně:
Je tvořena dvěmi zásadními systémy:
- nervovou buňkou – neuronem
- podpůrnými neurogliovými buňkami s jejich výběžky – jsou čtyři typy: 3 typy jsou ze zevního listu – ependymové buňky, astrocyty (astroglie a oligodendrocyty (oligodendrogile) Obrázek č. 1 Obrázek č. 1a a 4. je mikroglie (mezoglie, Hortegova glie), která se objevuje v nervové tkáni s prorůstáním cév – mesodermu. Funkce podpůrných buněk neurogliových je :
- podpůrná mechanická
- účast na myelinogenezi
- účast na degeneraci, regeneraci a růstu
- nutritivní úloha,
- účast na exkreci,
- účast na signalizaci mezi neurony a glií.
A k tomu i cévním systémem.
Nervová buňka – neuron Obrázek č. 2 Obrázek č. 3 Obrázek č. 4
Je základní morfologickou, trofickou a funkční jednotkou nervové tkáně. Odhaduje se jich asi na 100 miliard v mozku.
Skládá se z:
- těla – perikaryon, neurocyt, pyrenofor má trofickou funkci a zajišťuje životnost celé ho neuronu a tím i jeho specifickou funkci. Tvoří jej buněčná membrána, velké jádro a cytoplazma s buněčnými organelami (mitochondrie, lyzosomy, charakteristický Golgiho aparát, endoplasmatické retikulum s ribozomy). Pro nervovou buňku je typická tzv. Nisslova substance – propojené cisterny endoplasmatického retikula, která zasahuje do dendritů, ale nikoli do axonu. Cytoskelet se skládá z neurotubulů (mikrotubulů) a zvlněných filament. U některých neuronů jsou v neuronu zrnka pigmentu, melaninu (např. substatia nigra), dále tukové kapky s příměsí žlutého pigmentu, tzv. multivezikulární tělíska a multilamelární útvary.
- výběžky:
- dostředivých cytoplasmatických výběžků – dendritů – vedou dostředivé vzruch do neuronu, často se bohatě větví s jejich povrch zvětšují vybíhající dendritické trny – spinae. V buněčné membráně je mnoho receptorů, které představují místa chemicky aktivovaných iontových kanálů. Jsou to bílkovinné útvary reagující na chemické signály z jiných neuronů (např. na míšním motoneuronu se jich odhaduje asi 10 000, z toho 20 % na buněčném, 80 % na denritech, které mají 40 % povrchu membrány nervové buňky. Dle stupně větvení se dělí na primární, sekundární a terciální dendrity. Jsou tvořeny buněčnou membránou a v odstupu do nich proniká Nisslova substance. Označují se jako protoplasmatické výběžky, protože mají podobnou skladbu jako neuroplasma těla.
- odstředivě vedoucí, z nervové buňky vystupující jeden výběžkem – neurit, jehož základem je tzv. osový válec – axon Obrázek č. 5 Obrázek č. 6 Obrázek č. 7. Ten začíná po odstupu z těla neuronu – tzv. odstupový kužel – funkčně tzv. iniciálním segmentem, kde vzniká vzruch, a vede podráždění ve formě vzruchu k dalšímu neuronu nebo výkonnému orgánu – efektoru. Je zakončen terminálním úsekem – telodendrion, který se větví a na koncích větvení jsou tzv. terminální knoflíky (boutons terminaux).
- zakončení výběžků – synapse.
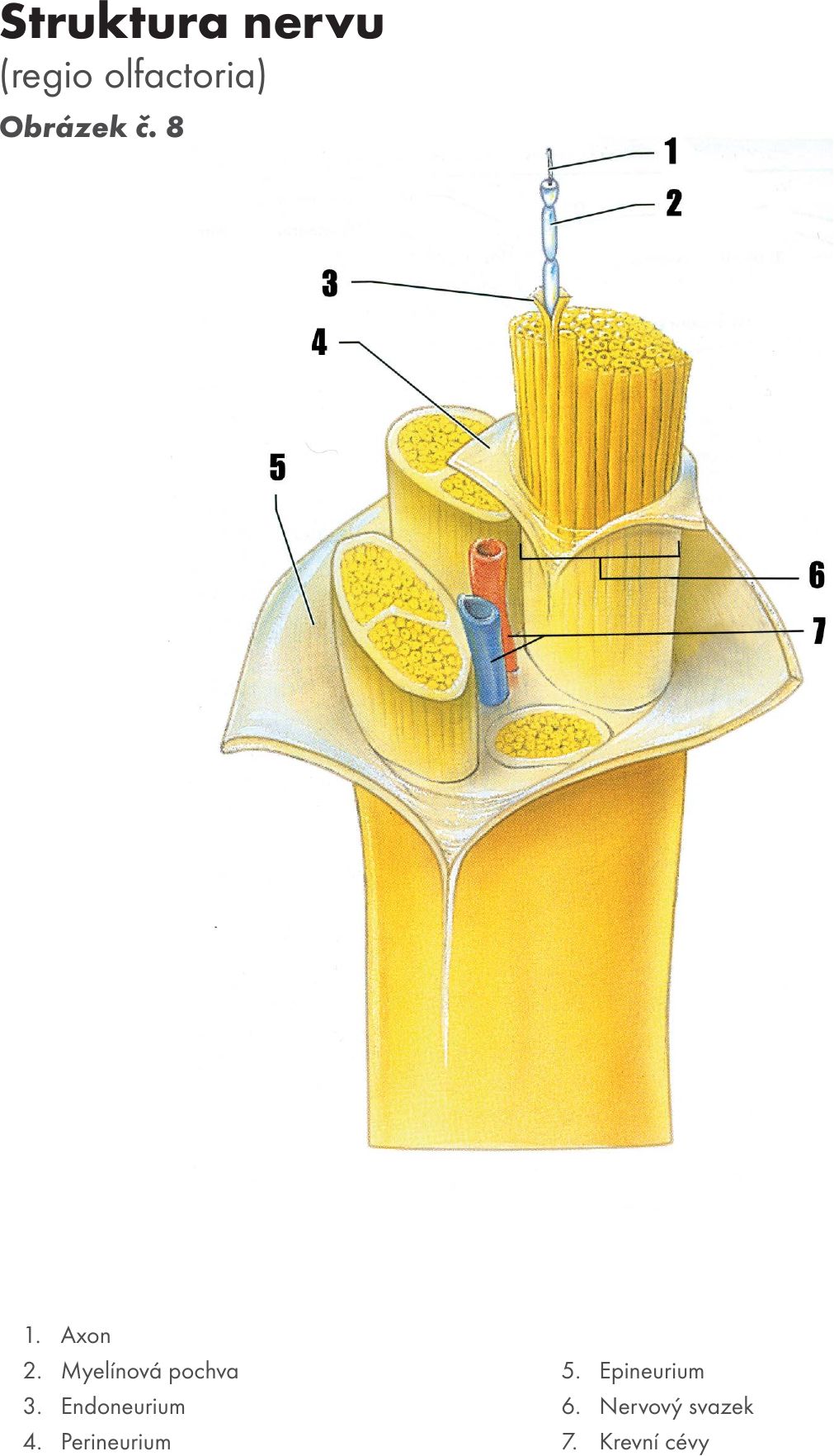
Skladba axonu Obrázek č. 8 Obrázek č. 9:
- povrchová membrána – axolema
- cytoplasmatická složka. Je tvořena cytoplasmou s neurofilamnenty a neurotubuly – mikrotubuly, mitochondriemi, nepravdelnými profily hladkého endoplasmatického retikula.
- obaly - dvou typů: myelinové (60 % v CNS) a bezmyelinové (40 % v CNS).
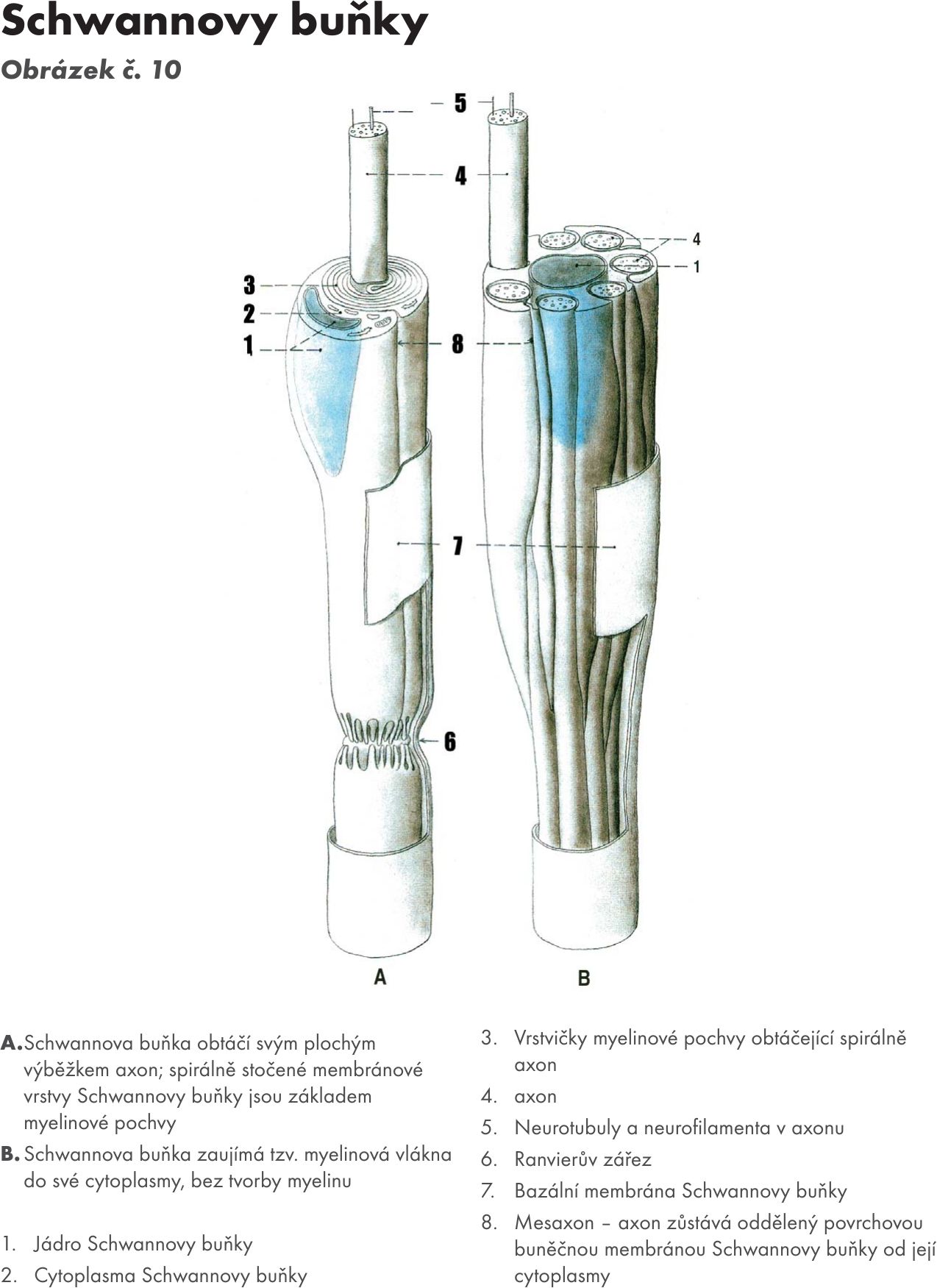
Myelinizované axóny (bílá a dřeňová vlákna, periferní nervstvo – PNS). Myelinovou pochvu tvoří v CNS z výběžků buněk oligodendroglie a v PNS z tzv. Schwannových buněk, které se postupně obtáčejí svou membránou i v několika vrstvách kolem axonu (až 50 vrstev, s výběžku zůstávají prakticky pouze membrány – dvoumembrána, cytoplasma mezi nimi se postupně ztrácí). Mezi buňkami se na axonu nacházejí tzv. Ranvierovy zářezy, kde není axon kryt Schwanovou buňkou, v tomto místě buňky pouze sousedí. Oblast krytá Schwanovou buňkou se nazývá internodální úsek. Na periferních nervech jsou v myelinové pochvě ještě zřetelné Schmidt-Lantermannovi zářezy, které odpovídají místům hranic mezi dvouvrstvami membrán Schwannových. V oblasti Ranvierova zářezu je axon ztenčelý a zde vydává i kolaterály. Myelin obsahuje 40 % proteinů a 60 % lipidů (fosfolipidy, cerbrozidy, sulfamidy a cholesterol). Vznik myelinu na axonech se označuje jako myelogeneze a myelinizace.
Poslední, třetí skupinu tvoří tzv. šedá Remakova vlákna (jsou to bezmyelinové axony vegetativních nervů), která jsou v podobě svazečku zahrnuta mezi jemné cytoplasmatické výběžky Schwannových buněk, bez toho aniž by se okolo axonu navrstvili lamely myelinu. Převážně se axony v podobě kruhovitých shluků vtlačují do periferní části cytoplasmy Schwannových buněk Obrázek č. 10.
Axoplasmatický transport je antegrádní – od těla axonu k zakončení a retrográdní, který je o polovinu pomalejší. Je to aktivní děj a ne pouhá difúze. Je potřeba k jeho činnosti energie (oxidační fosforylace, ATP, optimální koncentrace vápníkových iontů – 3–5 mmol). Podílejí se na něm tzv. transportní filamenta.
Dělí se na:
- pomalý transport (pod 1 mm až několik mm za 24 h) – dopravuje 80 % rozpustných bílkovin, které se během transportu rozkládají a jsou významné pro funkci axonu
- rychlý transport (40–700 mm / 24 h, optimum cca 400 mm /24 h) – dopravuje především některé neurotransmitery (např.noradrenalin) enzymy jejich přeměny (např. dopamin-β- hydroláza, acetylcholinesteráza), aminokyseliny, 20 % bílkovin a glykoproteidy, zejména některé důležité složky membrány neuronu, mezi nimi i receptory. Uvolňují se buď okamžitě, nebo postupně – ty se označují jako látky se středně rychlým transportem.
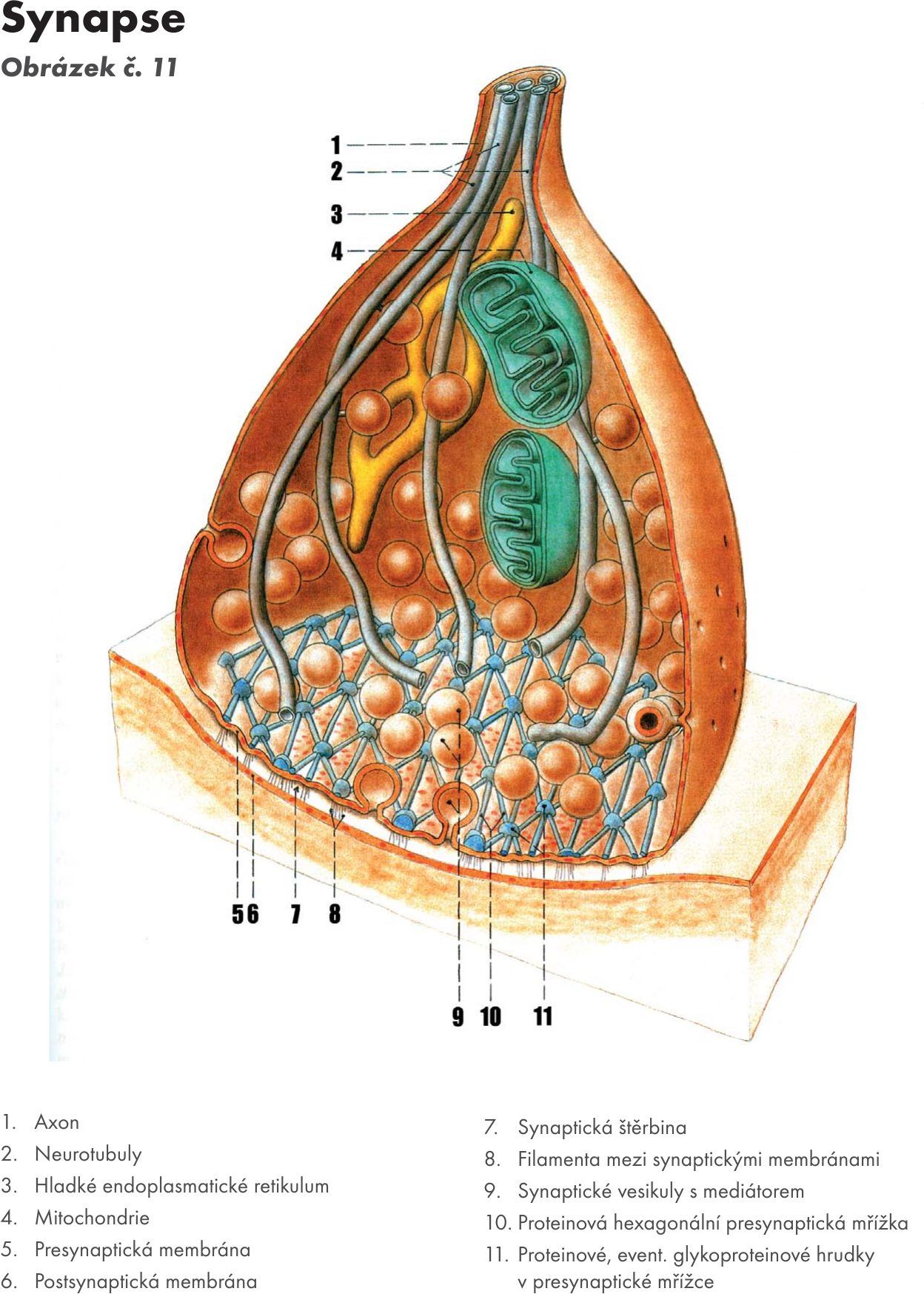
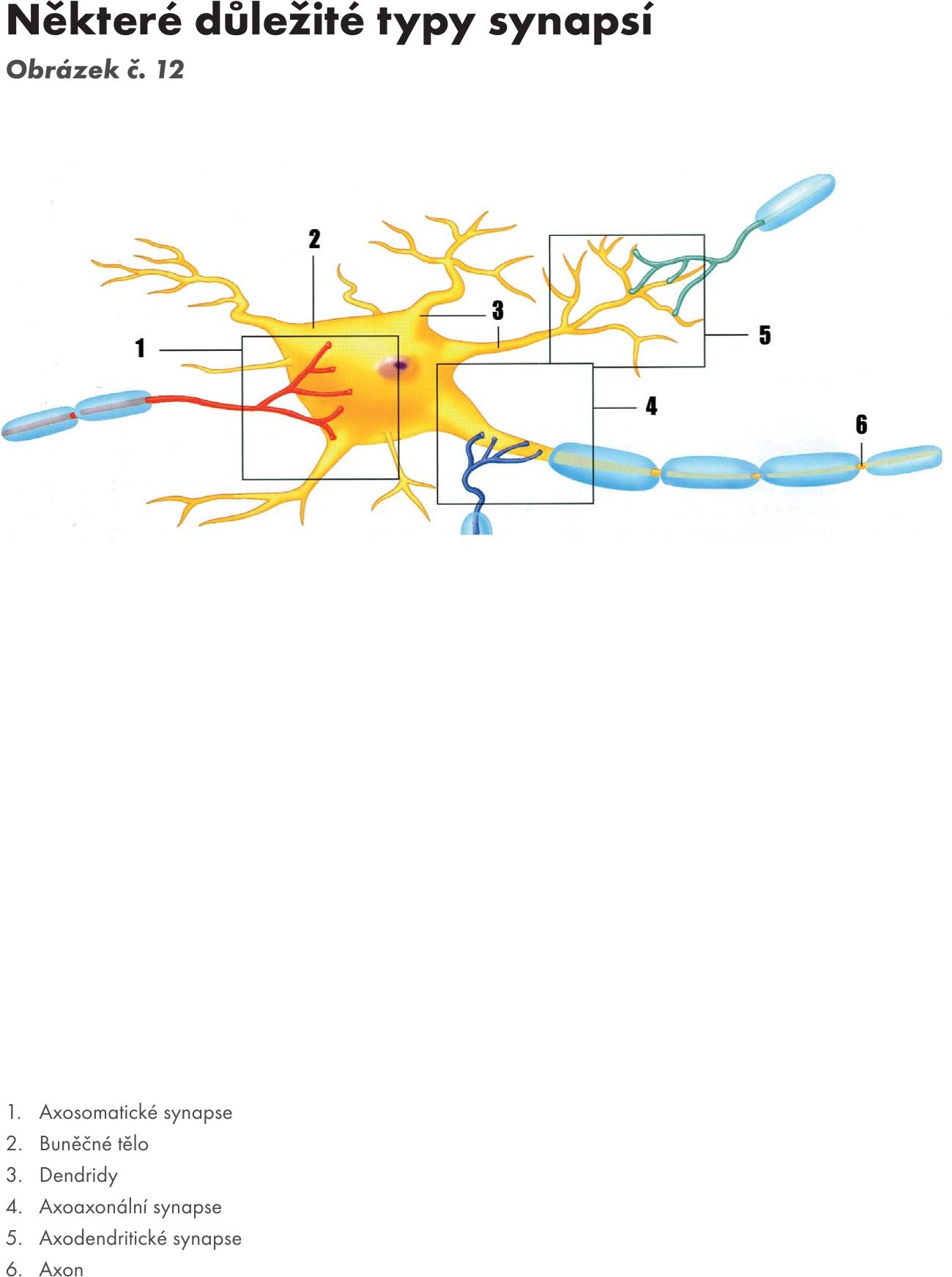
Synapse – zápoje Obrázek č. 11 Obrázek č. 12. Specifické útvary zajišťující funkční spojení mezi neurony. Jeden neuron je presynaptický a druhý postsynaptický, což zaručuje jednosměrnost vedení podráždění v nervovém systému.
Synapse axonu. Nazývají se také jako transmisivní synpatický segment. Je vytvořen rozšířenými zakončeními bez myelinu. Může jich být u jednoho axonu až 100. Označuje se jako presynaptická část synapse. Obsahuje mitochondrie a látky dopravované axoplazmatických transportem. Obsahuje presynaptické váčky, které obsahují zásobu mediátorů – transmiterů.
Klasické schéma : presynpatický neuron (axon → terminální synaptické zakončení) → dendrity nebo soma postsynaptického neuronu. Přenos se mezi pre- a postsynaptickým neuronem uskutečňuje humorálně. Cytoplasma presynaptických zakončení obsahuje vedle mitochondrií (zdroj energie) synaptické váčky (vehikuly), které se hromadí především u synaptické štěrbiny (cca 20nm) a obsahují neurotransmiter (transmiter, přenašeč, posel, mediátor). Přes presynaptickou membránu, která se vyznačuje presynaptickým zahuštěním (denzitou) přechází neurotransmiter do synaptické štěrbiny, a pak dochází ke vzniku podnětu na postsynaptické membráně, a to především v části označované jako subsynaptická membrána, kde je rovněž zahuštění – postsynaptická denzita a pod touto částí membrány je uložen subsynaptický aparát.
Buněční receptory jsou útvary vyčnívající z vnější strany membrány, aby byla umožněna vazba na mediátoru na aktivní vazebné místo receptorové bílkoviny. Jedná se převážně o specifické vazby.
Jsou dvě skupiny receptorů:
- chemicky řízené iontové kanály (např. acetylcholin nikotinový receptor – Na+ a K+ Ca++, receptory pro útlumové kyseliny propouštějící Cl-) – mění ionotovou vodivost.
- neměnící iontovu vodivost – biochemické reakce neuronu prostřednictvím druhých buněčných poslů (cyklické kyseliny adenylfosforečná, guanidylfosforečná, lipidové látkym např diacylglycerol apod.).
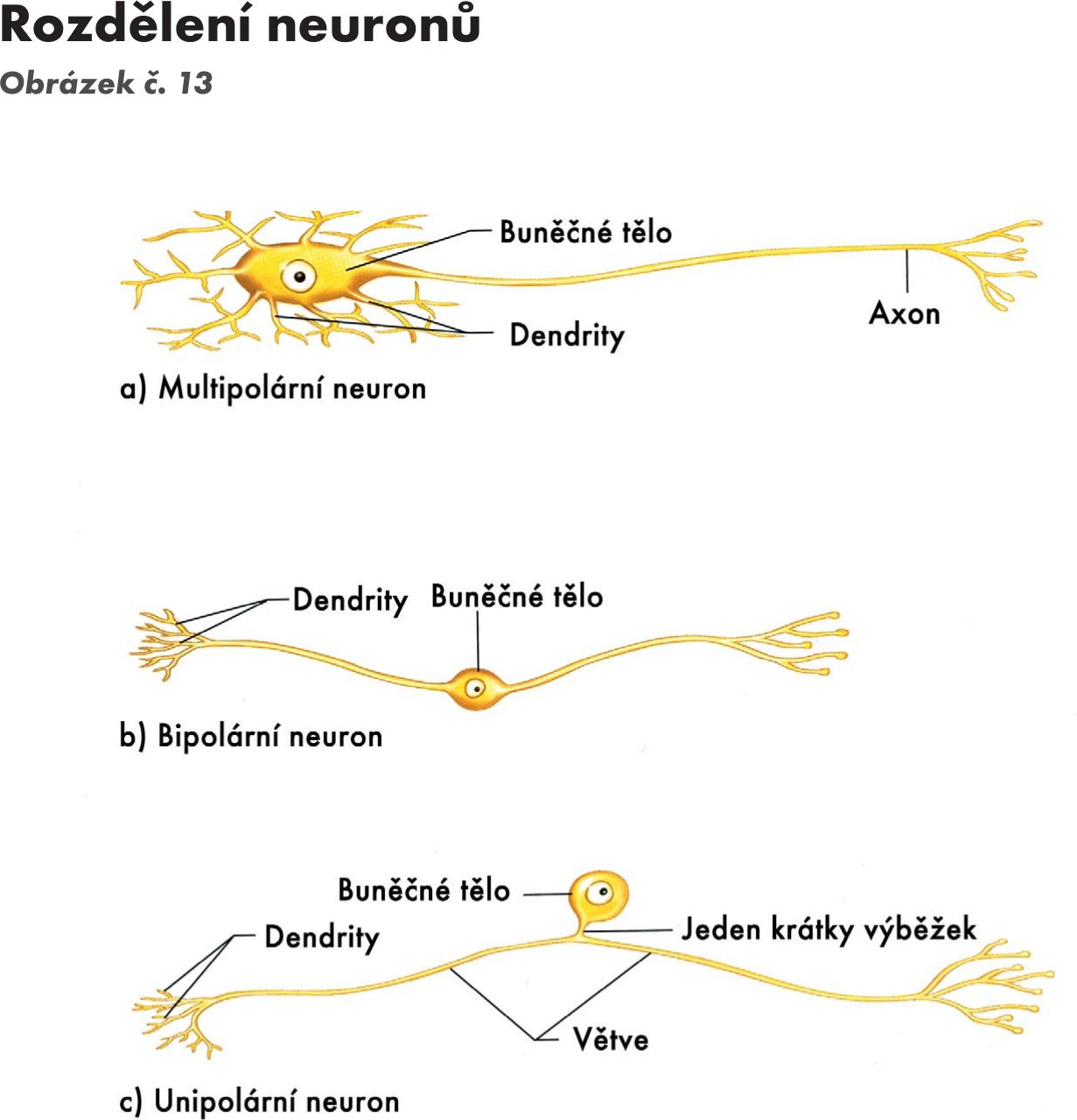
Nervové buňky se dělí na Obrázek č. 13:
- Multipolární - mají všechny části, mají více dendritů.
- Bipolární - mají jeden dendrit a jeden axon – senzorické buňky – mají jeden relativně dlouhý axon (v sítnici, v předsíňovém gangliu,.
- Unipolární - mají jeden výběžek (tyčinky a čípky), podskupinou jsou pseudounipolární buňky vznikají přeměnou bipolární buňky, kde výběžky splynou v jeden nevelký vystupující z buňky, který se pak dělí ve tvaru písmene T.
- Apolární - nemají výběžky (v době histogeneze, přetrvávají jako receptorové buňky polohově sluchové a chuťové).
Dle ramifikace dendritů se dělí na :
- Izodendritické neurony – mají dlouhé přímočaře v jedné rovině orientované dendrity, po odstupu z těla se málo větví, ale vzniklé větve jsou delší než mateřská (v retikulární formaci mozkového kmene).
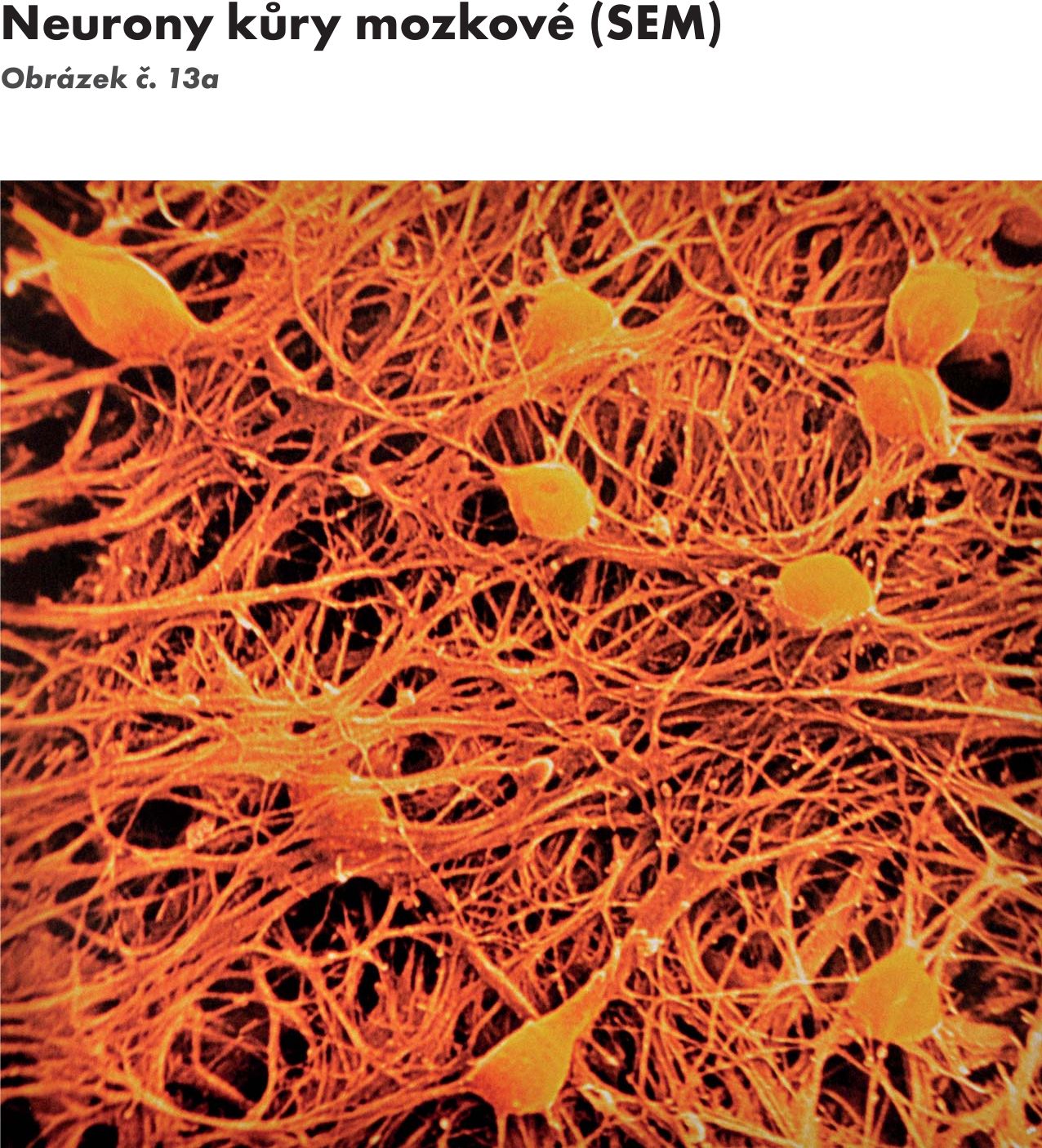
- Alodendritické neurony – mají několik hlavních větvících se kmenových dendritů (pyramidové buňky mozkové kůry a přepojovací jádra v mozkovém kmeni). Obrázek č. 13a
- Idiodendritické neurony – mají stromečkovitě se větvící dendrity a patří do skupiny specializovaných neuronů (mitrální buňky čichové kole, bulbus olfactorius, bipolární buňky v sítnici, v ganglion spirale cochlae, Purkyňové buňky mozečku…).
Wallerova degenerace, regenerace.
Wallerova degenerace vznikne po oddělení axonu od buněčného těla a dojde k degeneraci distálního pahýlu (tzv. retrográdní, sekundární – je časově kratší) a později i k anterográdní degeneraci (primární Wallerova degenerace – je časově delší a pozdější), kdy dojde, pokud nedojde k regeneraci a ke kontaktu s receptorem či efektorem periferie axonu, i k postupnému zániku těla neuronu (např. příčina centralizace bolesti).
Wallerova regenerace je reakce těla buňky na oddělení těla s produkcí stavebních proteinů od axonu. Nový axon roste rychlostí cca 1 mm/den a snaží se spojit s původními efektory nebo receptory, poté dochází k tvorbě myelinových pochev a následně k i event funkční regeneraci. Pokud je přerušen periferní nerv a nedojde k jeho spojení vznikne na jeho konci tzv. amputační neurom, který je citlivý na poklep.
Základní funkční projevy nervového systému
Ústřední nervstvo je soubor nejvyšších center, které udržují jednotu organismu a zabezpečují jeho integraci s prostředím, včetně reakcí. Množství zpráv a vlivů z vnějšího i vnitřního prostředí organismu se označují jako podnět. Nervový systém pak řídí, reguluje a kontroluje všechny funkce organismu a dává vznik k složitému komplexu činností, které označuje jako chování jednotlivce. Nervový systém má kódy, kterými získává podněty z prostředí (vnějšího i vnitřního), zpracovává informace a přiměřeně reaguje. Kódy v nervovém systému jsou v podstatě dvojí povahy : elektrické a humorální, které se mezi sebou vzájemně ovlivňují a integrují.
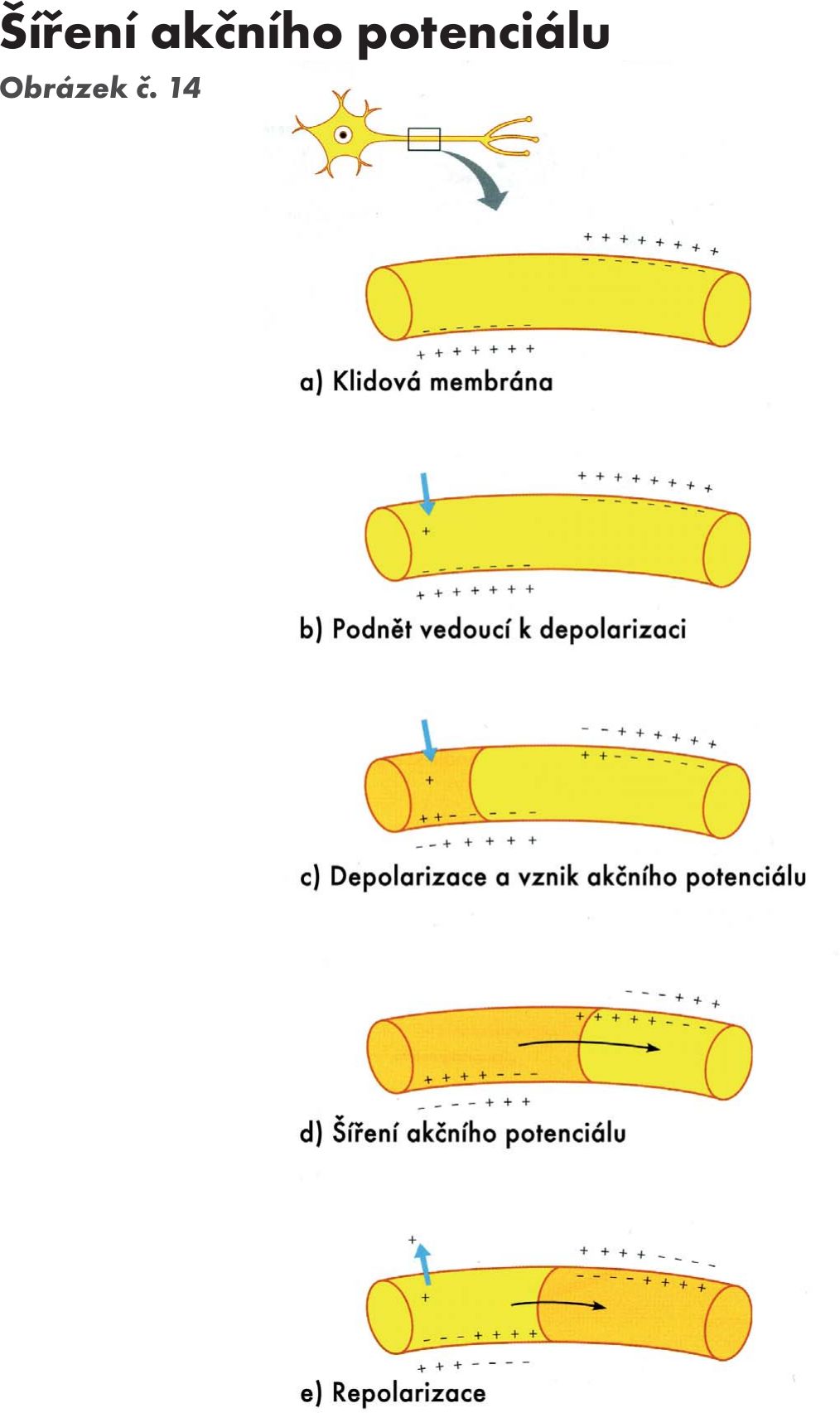
Základními ději v nervové systému jsou podráždění a útlum (excitace a inhibice) Obrázek č. 14. Obě děje jsou aktivní a vyžadují energii. Základem je změna propustnosti Na+ a K+ kanálů závislé na polarizaci membrány. Depolazirace subsynaptické membrány nestačí na aktivaci celé membrány – vznikne tak potenciálová změna, která se označuje jako excitační postsynaptický potenciál (EPSP) (depolarizace nastane cca 0,5 ms po dosažení aferentního vzruchu k tělu neuronu, za 1–1,5 ms dosáhne vrcholu, a pak se exponenciálně vrací k výchozí hodnotě (1/e tj. 1/ 2,718 se označuje časová konstanta, u ESPS je 4 ms). Postupně dochází k sumaci jednotlivých EPSP a dosáhne úrovně vzruchové aktivity – vzniká hrotový akční potenciál. Sumace proběhne současně a nazývá se prostorová sumace. Pokud podněty přichází v kratším intervalu než je depolarizace membrány, jedná se o časovou sumaci. Oba typy sumací se nazývají facilitace (odpověď převyšuje součet jednotlivých reakcí). Přechod vzruchu přes synapsi vyžaduje určitý časový úsek a označuje se jako synaptické zpoždění (je asi do 0,5 ms). Při depolarizaci dochází k přechodu sodíkových iontů dle koncentračního elektrického gradientu do neuronu. Podráždění některých vláken může způsobit hyperpolarizaci (vzniká asi za 1–1,25 ms po příchozu aferetního vzruchu), snižuje se dráždivost neuronu, tlumí se odpověď – inhibiční postsynaptický postenciál (IPSP), jehož vrchol nastává za 1,5–2 ms, klesá s časovou konstantou kolem 3 ms. I zde existuje prostorová a časová sumace, dochází k lokálnímu zvýšení propustnosti pro draslíkové a chlorové ionty, ale propustnost pro sodíkové ionty zůstává nezměněna. Výstup chlorového iontu z buňky je aktivní děj.
Principy kódování v nervstvu. Podněty dělíme dle tří kriterií: modality (specificity), kvantity a kvality. Pro vznik podnětu jsou nutné tří podmínky: musí být dostatečně silný (podnětový práh), musí mít určité minimální trvání, rozdíl mezi výchozím a novým stavem musí nastat dostatečně rychle. Při splnění podmínek se vybaví odpověď – vzruch dle zákona vše nebo nic.
Další důležité prvky v nervovém systému jsou : princip redundance, který je základním předpokladem plasticity.
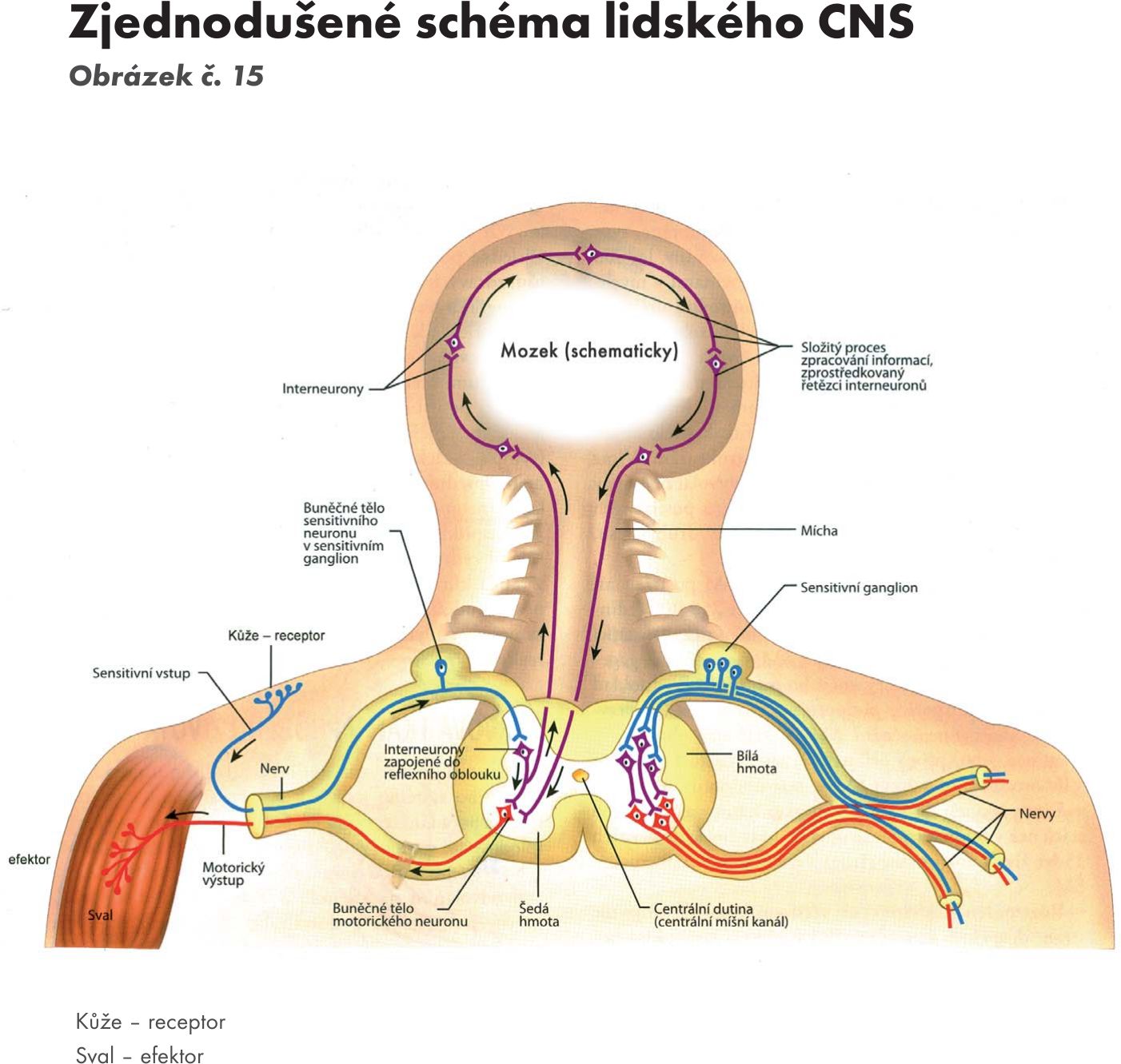
Reflex Obrázek č. 15 (autor pojmu René Descartes) je funkčně strukturní jednotka nervového systému, v níž se projevují podráždění a útlum. Reflex se uskutečňuje po reflexním oblouku, který tvoří: čidlo – receptor → dostředivá (aferentní) senzitivní dráha → ústředí v nervovém systému – v míše, mozku → odstředivá (eferentní) motorická dráha → efektor – sval, žláza. Dělí se dle počtu neuronů : dvouneuronové – propriocepční, tříneuronové, disynaptické – exterocepční, víceneronové – vegetativní, viscerální. Dále se dělí na nepodmíněné – vrozené a podmíněné – získané, naučené. Zvláštní skupinou jsou instinkty a tzv. patologické reflexy neboli starší obranné reflexy, které jsou překryty či potlačeny nadřazenými centry a projevují se po jejich vyřazení.
Základní funkční výkony nervových center jsou prostorová sumace – součet podnětů a okluze – jedna buňka podrážděna současně z více vláken, podnět je slabší než pokud by byla podráždění izolovaná, facilitace – zvýšené sumační podráždění. Při základních dějích v centrálním nervstvu, podráždění a útlum, vzniká v okolí míst podráždění útlum a v okolí místa útlumu podráždění – prostorová indukce. Pokud vznikne opačný děj po skončení děje prvotního – indukce časová. Indukce jsou typické pro vyšší nervové činnosti. Další jsou revertebrace – tj. kroužení vzruchu po neuronových okruzích a vznik tzv. následného výboje (after-discharges). Revertebrace je podkladem tzv. rozněcování (kindling), je považována za obecný základ fyziologických a patologických pochodů (např. epilepsie).
Uspořádání nervového systému. Prostorová síť neuronů a jejich spojů představuje složitý komplex obvodů. Obecnými pravidly funkčně strukturního uspořádání jsou: princip konvergence – sbíhání se ke koncovému neuronu, princip divergence – větvení paralelní i seriové, princip zpětné vazby.
Základní kompenzační strategií nervového systému je plasticita nervového systému. Je to schopnost přizpůsobovat se nejrůznějším vlivům. Důležité prvky kompenzační strategie jsou: redundance – princip nadbytečnosti, alternace – přejímání funkce, vikarující zastupující funkce – přebírání úlohy vyřazené části jinými fyziologickými mechanismy, diaschiza – funkce je vyřazena pro nedostatek přítoku vzruchů nebo naopak nadměrnou impulsací, a tím vzniká útlum.
Struktura periferního nervu:
Periferní nervová vlákna mají tendenci se sdružovat do svazků – nervů. Jednotlivá vlákna nervových buněk jsou zabalena do ochranného pláště, endoneuria. Více neuritů vytváří primární svazky a jsou obaleny v dalším ochranném plášti, jímž je perineurium, které vypadá jako rukáv tvořený vrstvami epiteloidních buněk. Buňky jsou uspořádány v několika vrstvách a navzájem jsou propojeny pomocí těsných spojení (tight junctions), které zajišťují nepropustnost pro většinu makromolekul. Tato vnitřní vrstva je pak obalena vnější vrstvou z pojivové tkáně. Posledním ochranným pláštěm je epineurium. Stavbu těchto periferních nervů lze tedy přirovnat ke stavbě svalového vlákna.16
16 Wikipedie: Otevřená encyklopedie: Perineurium [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Perineurium&oldid=10407176
Každý axon neuronu je izolován Schwannovou buňkou, okolo níž se nalézá první ochranný obal endoneurium. Endoneurium je vrstvička z pojivové tkáně okolo každého nervového vlákna periferních nervů a je tvořeno vrstvou retikulárních a kolagenních vláken, dále se zde nalézají fibroblasty a příležitostné makrofágy. Několik svazků je chráněno druhým obalem perineuriem, tvořeným z epiteloidních buněk. Perineurium je ochranný plášť svazku několika nervových vláken, která jsou jednotlivě obalena ve svém vlastním pouzdře zvaném endoneurium. Více svazků perineuriích je pak obaleno dalším pláštěm nazvaném epineurium. Posledním obalem je epineurium, které chrání a zároveň vyplňuje svazky vláken.17 Epineurium je tvořeno fibrózním pláštěm vazivové tkáně. Celý útvar se pak nazývá periferní nerv. Vazivová tkáň obklopující nerv se nazývá mezoneurium. Důležitou složkou mezoneuria jsou výživné cévy (vasa nervorum) a lymfatické prostory.
17 Wikipedie: Otevřená encyklopedie: Epineurium [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Epineurium&oldid=10144339
V podélném směru jsou atonální vlákna zvlněná – tzv. Fontanova undulace, což umožňuje nervům protažení v podélném směru a brání trakčnímu poranění při běžném pohybu pohybu.
| Obrázky ke kapitole: Základní pojmy | |||
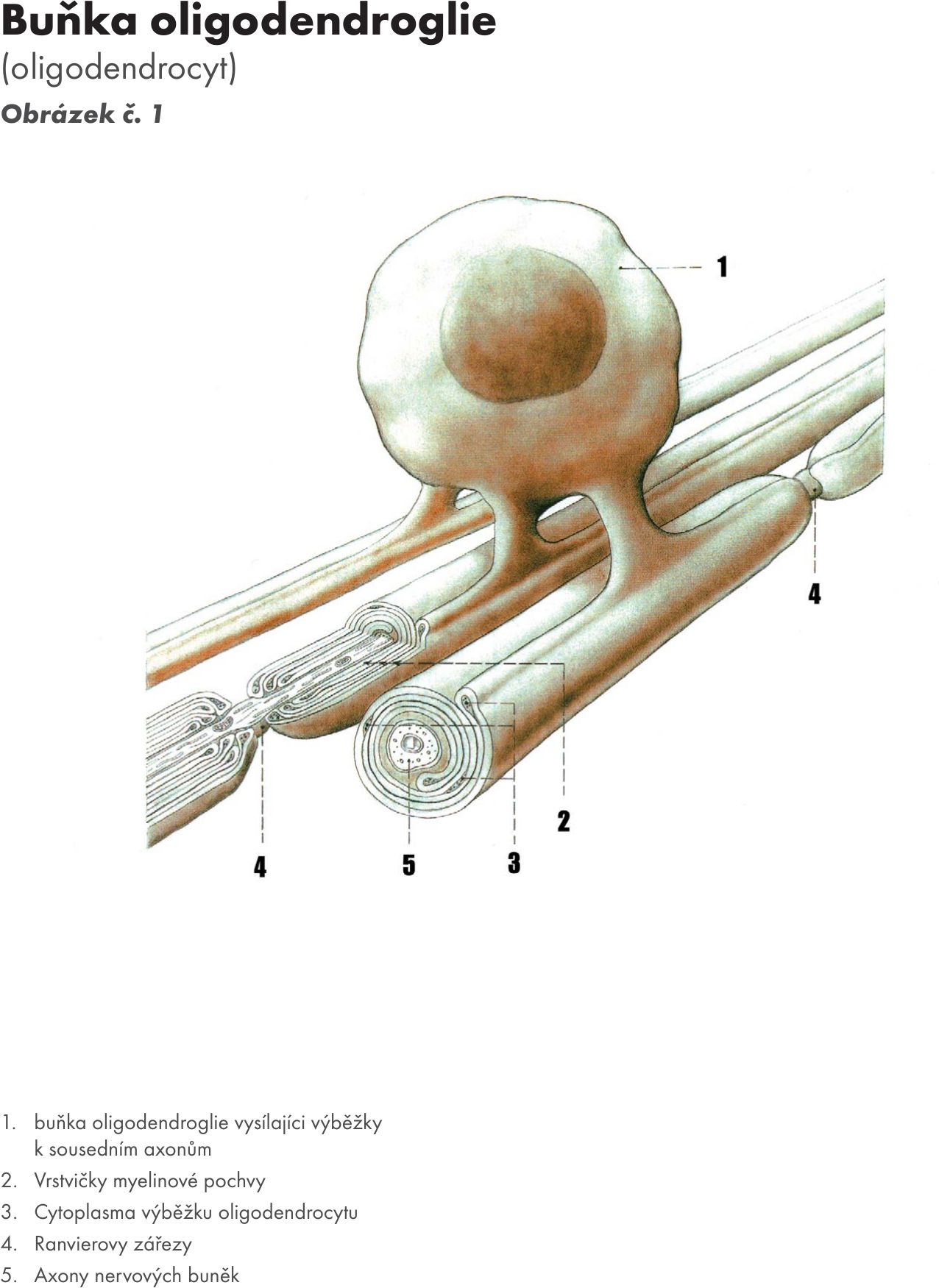
Obr. 1 Buňka oligodendroglie
|
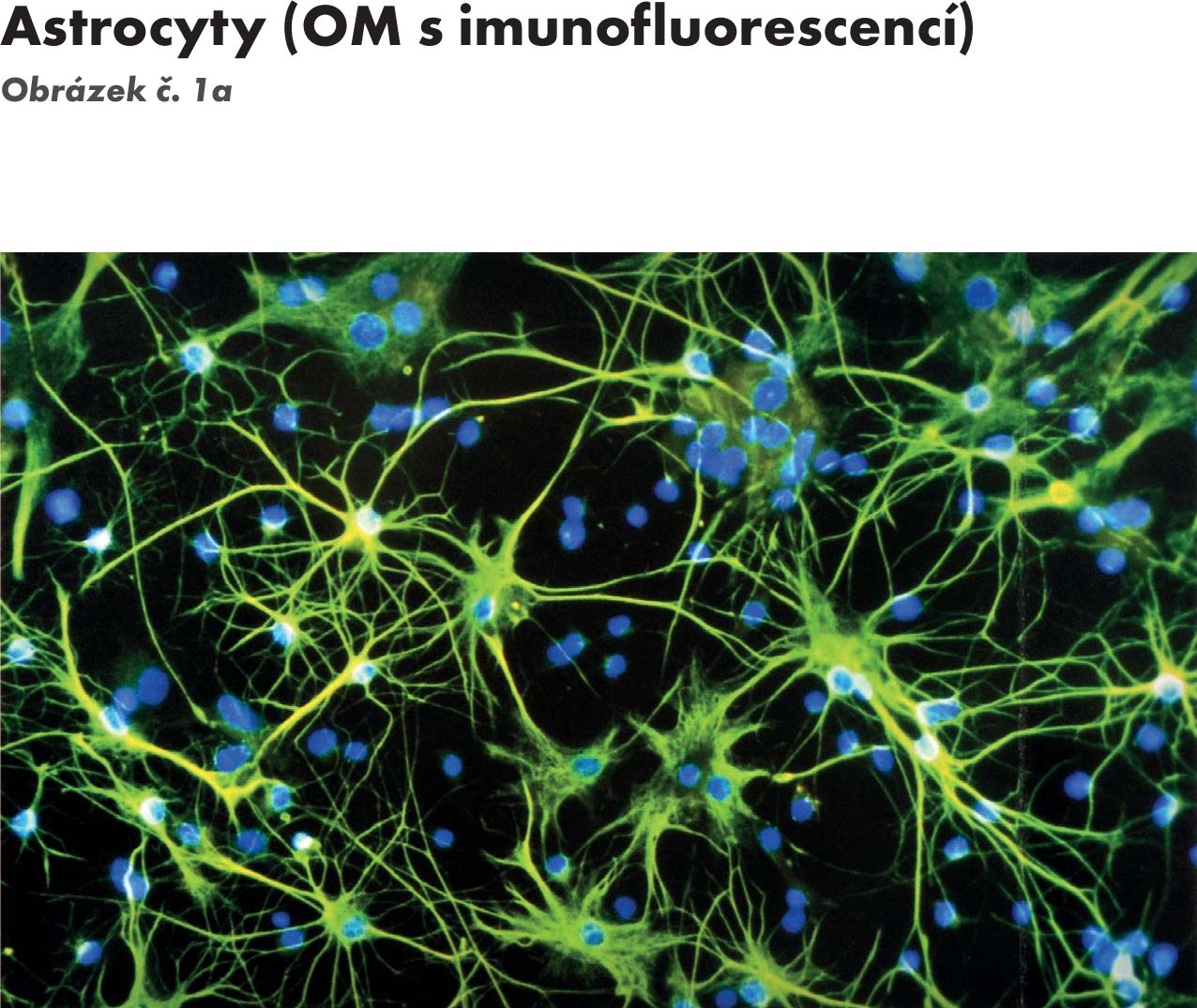
Obr. 1a Astrocyty (OM s imunofluorescencí)
|
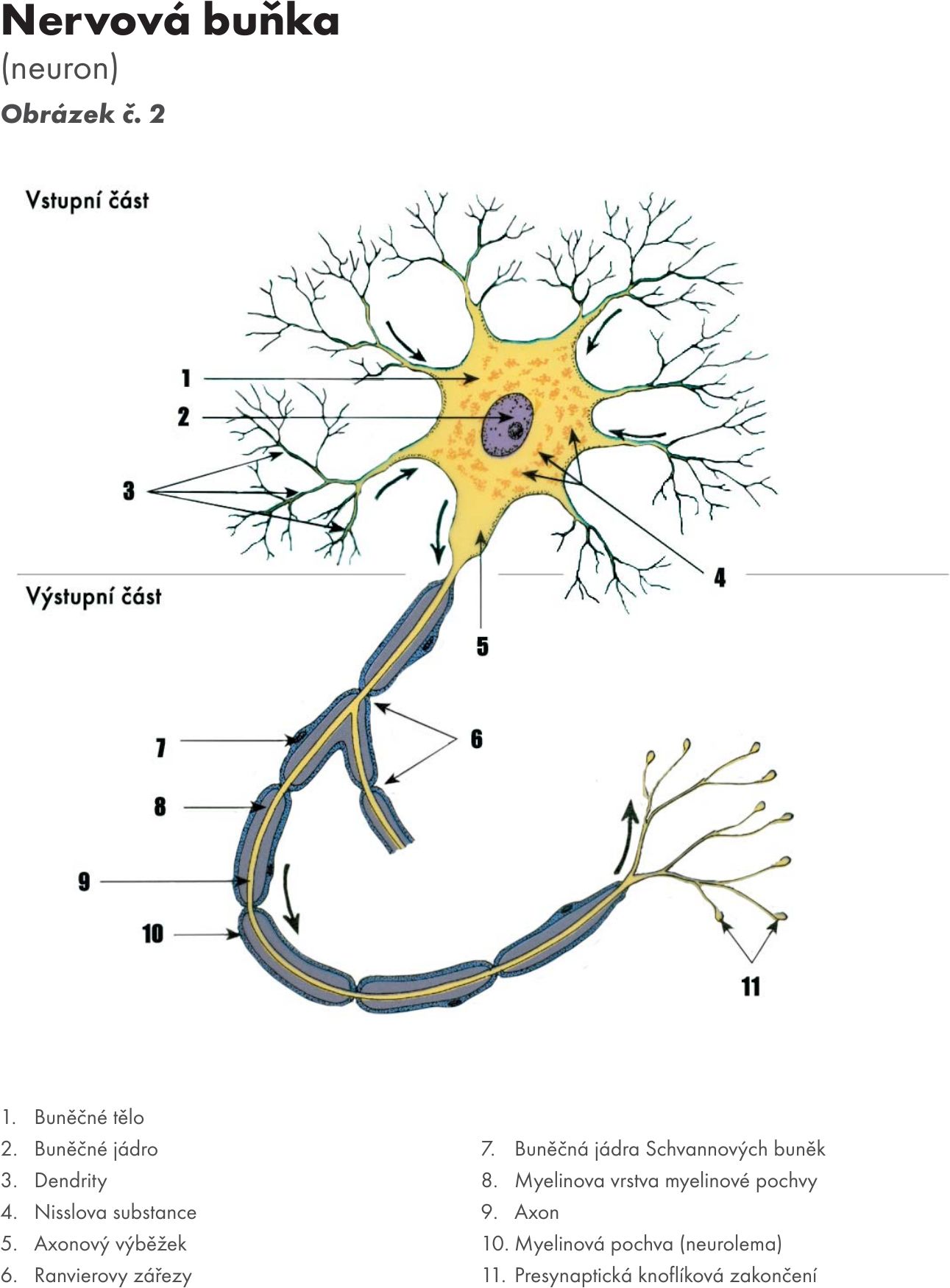
Obr. 2 Nervová buňka
|
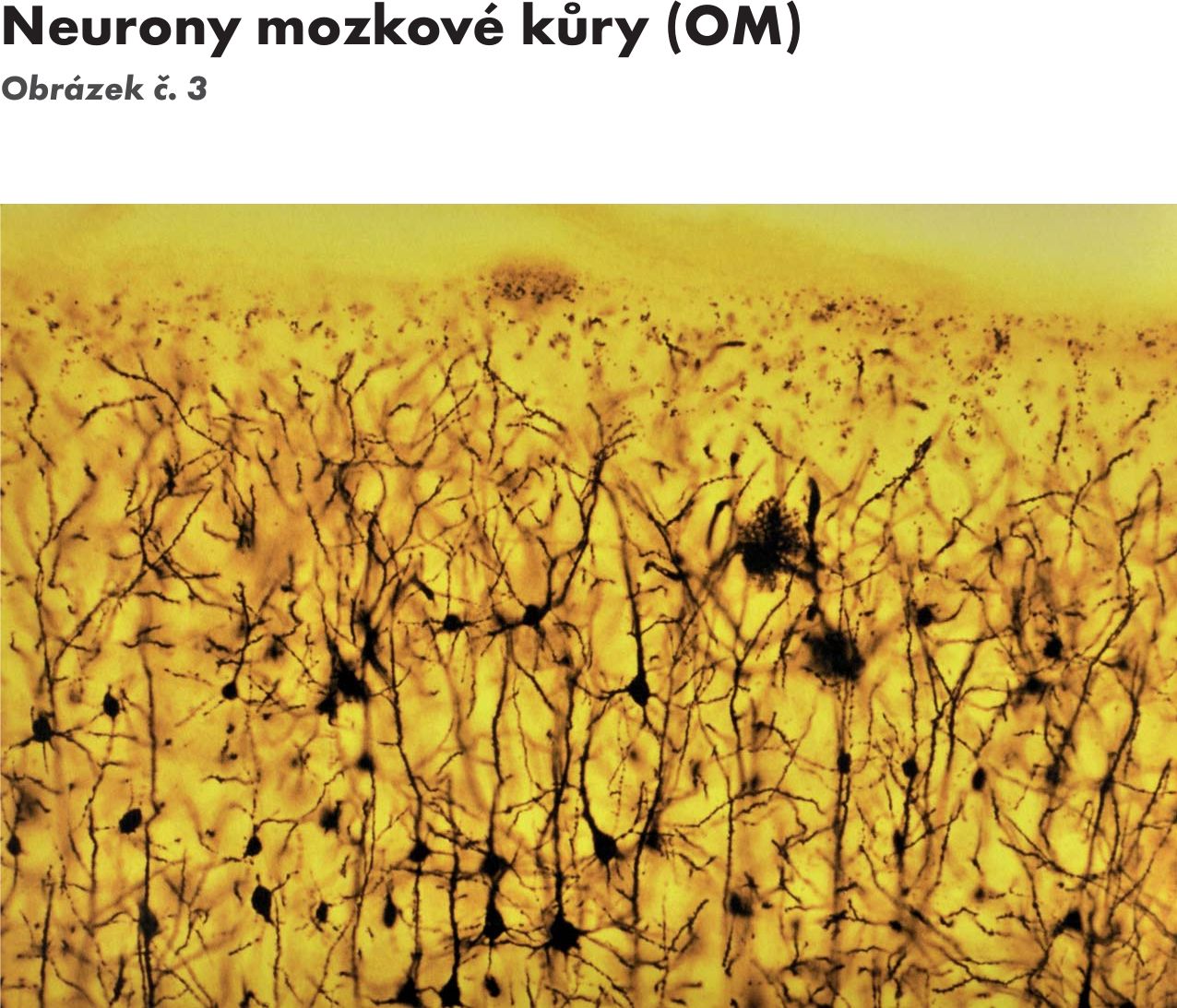
Obr. 3 Neurony mozkové kůry (OM)
|
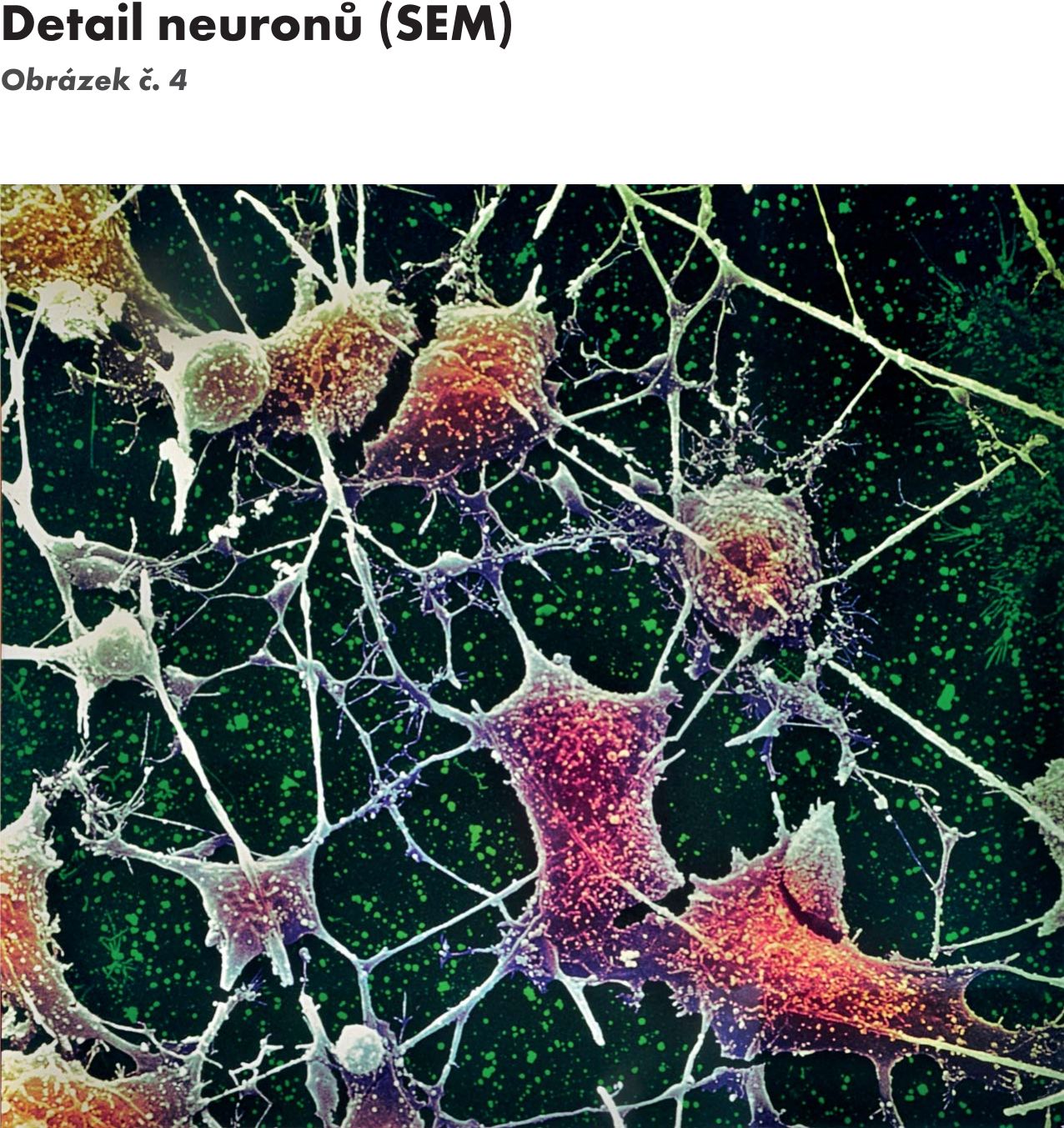
Obr. 4 Detail neuronů (SEM)
|
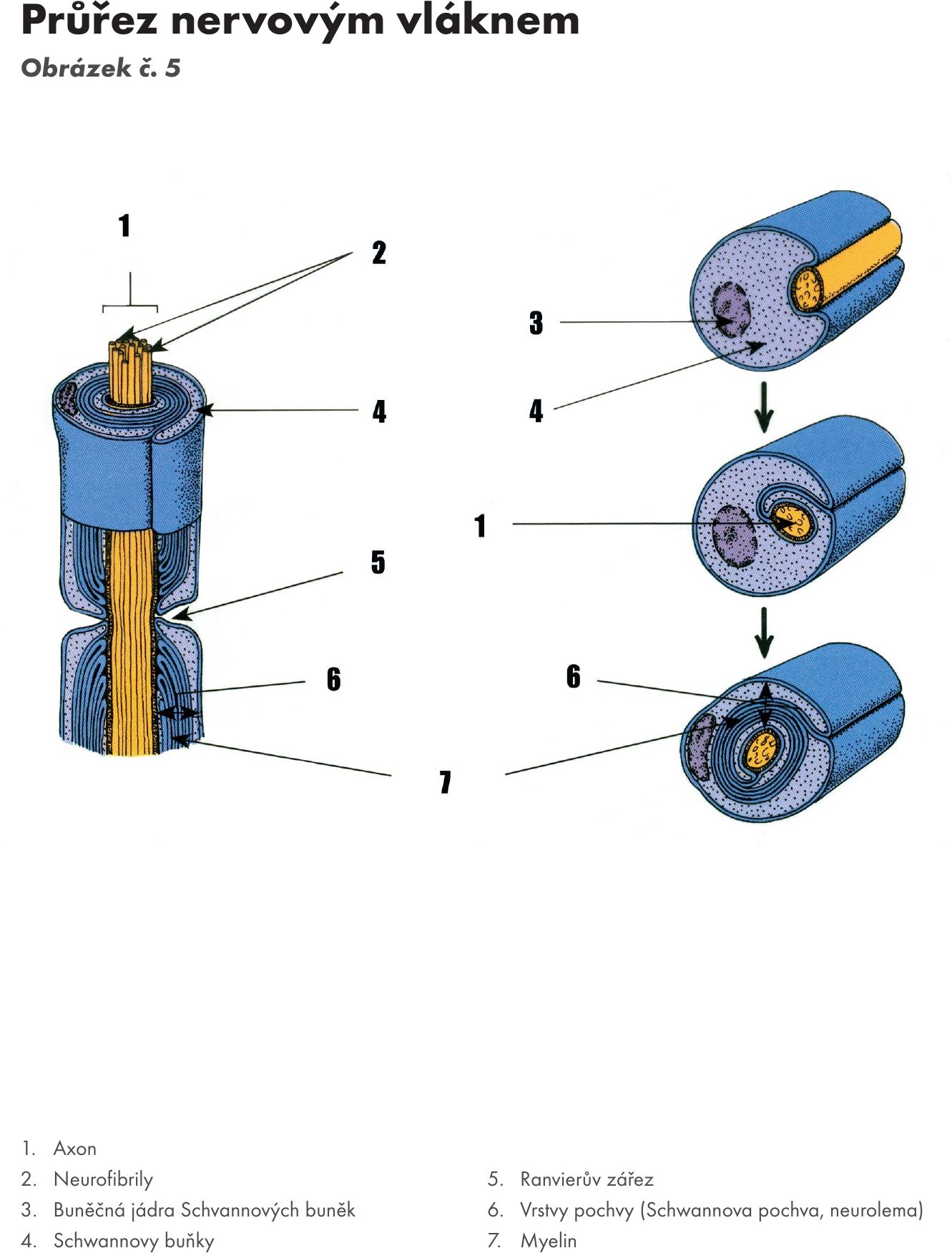
Obr. 5 Průřez nervovým vláknem
|
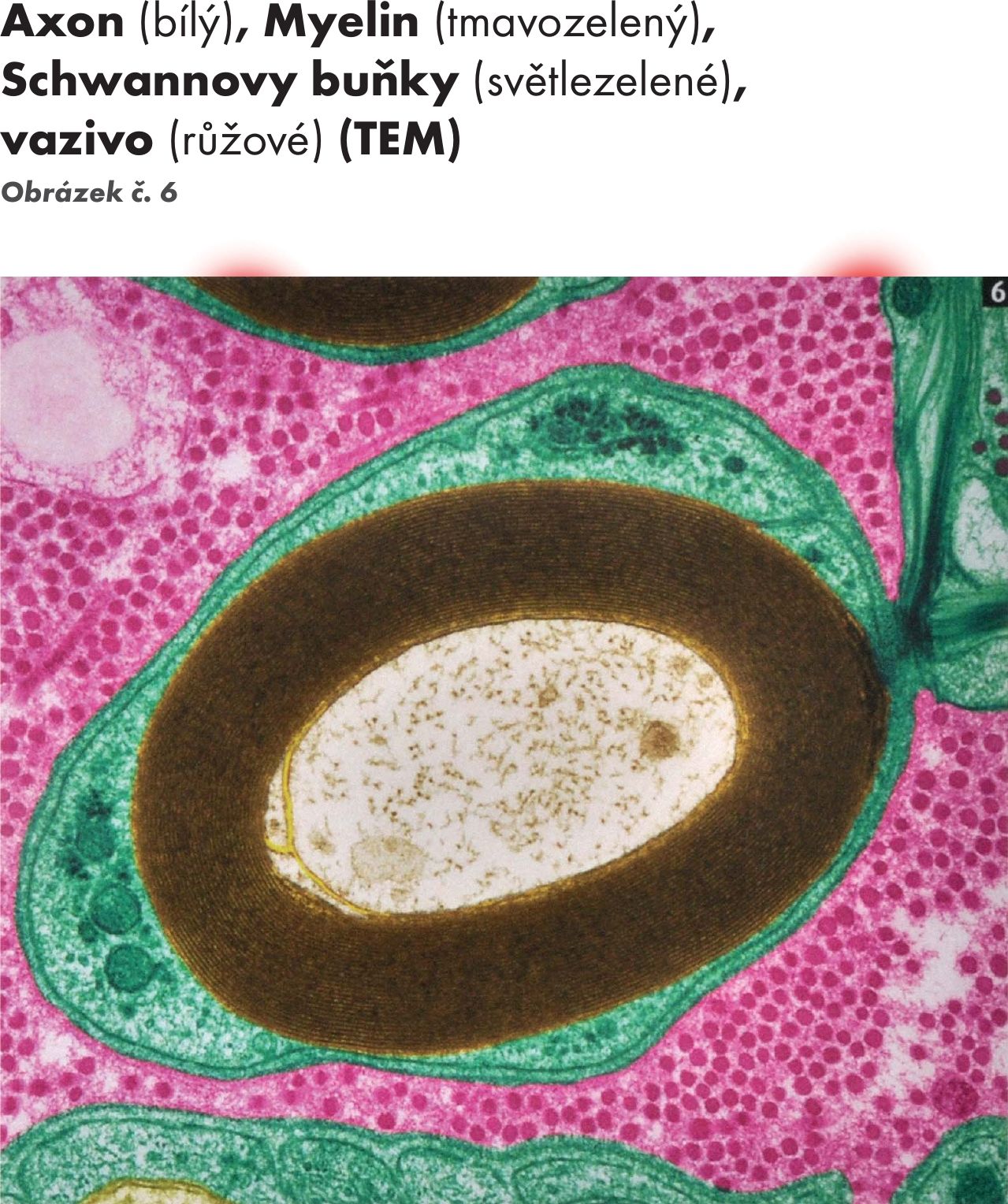
Obr. 6 Axon, Myelin, Schwannovy buňky, vazivo (TEM)
|
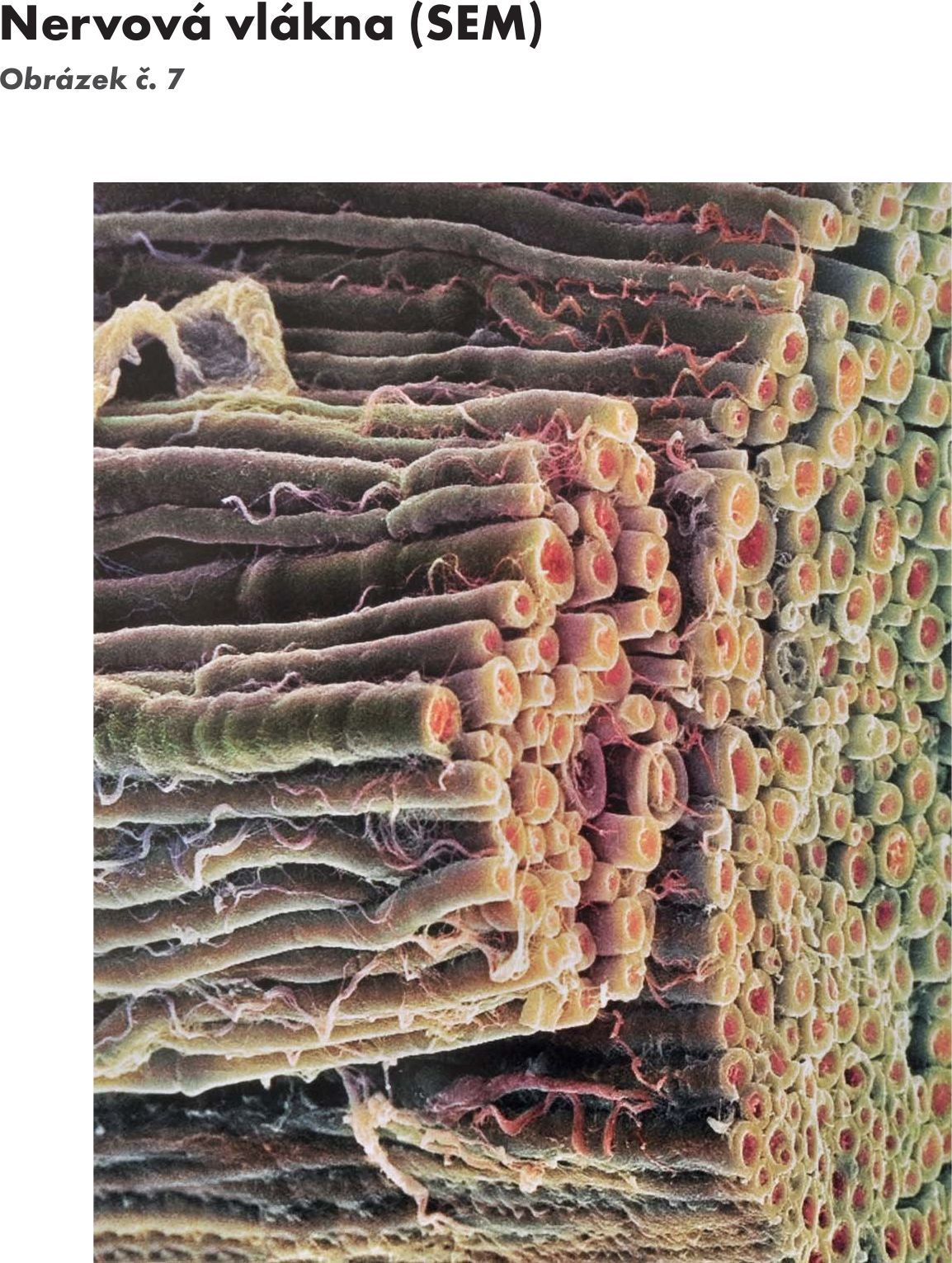
Obr. 7 Nervová vlákna (SEM)
|
Obr. 8 Struktura nervu
|

Obr. 9 Průřez nervem
|
Obr. 10 Schwannovy buňky
|
Obr. 11 Synapse
|
Obr. 12 Některé důležité typy synapsí
|
Obr. 13 Rozdělení neuronů
|
Obr. 13a Neurony kůry mozkové (SEM)
|
Obr. 14 Šíření akčního potenciálu
|
Obr. 15 Zjednodušené schéma lidského CNS
|
|||
Hřbetní mícha – medulla spinalis
Hřbetní mícha je provazec nervové tkáně uložený v páteřním kanálu délky asi 40–50 cm Obrázek č. 16. Na průřezu v různých segmentech má oválný, kruhovitý nebo válcovitý tvar o šířce 10–13 cm. Je obklopena míšními obaly. Jsou tři: na povrchu je měkká plena- pia mater spinalis, která vniká do všech záhybů a rýh míchy. Nad ní je pavoučnice – arachnoidea spinalis, která míchu obaluje volně. Mezi oběma měkkými plenami je prostor – cavitas subarachnoidea vyplněný mozkomíšním mokem – liquor cerebrospinalis. Zevně je tvrdá plena- dura mater spinalis, která kolem míchy vytváří vak – saccus durae matris spinalis a přechází až na míšní nervy do meziobratlových otvorů – foramina intervertebralia. Tvrdá plena nesrůstá s periostem kanálu páteřního.Mícha začíná pod velkým týlním otvorem - foramen magnum výstupen prvního krčního míšního nervu. V tomto místě se také kříží motorické dráhy – decussatio pyramidum.
Kaudálně mícha končí kuželovitě v úrovni L1–2 – conus medullaris a dále pokračuje do úrovně S2 konečným vláknem – filum terminale. Na míše se nachází dvě ztluštění – krční, má střed ve výši obratle C5, a bederní, ve výši obratle Th12 (intumescentia cervicalis et lumbalis). Jsou to místa odstupu pažní a bederní pleteně pro inervaci horních a dolních končetin.
Zevní popis:
Štěrbinou a brázdami je povrch míchy podélně rozdělen na přední, boční a zadní provazce – funiculi medullae spinalis ventralis, lateralis et dorsalis. Zadní boční brázdou – sulcus dorsolateralis vstupují vlákna zadních kořenů míšních nervů (senzitivní – radices posterior). Na zadních kořenech jsou nervové uzliny vřetenovitého tvaru (ganglia), která obsahují pseudounipolární buňky.
Přední boční brázdou – sulcus ventrolateralis vystupují vlákna předních kořenů míšních nervů, motorická – radices anteriores. Přední a zadní kořeny se spojují v míšní nervy, kterých je 31–33 párů a vystupují z meziobratlových otvorů. Z toho je 8 nervů krčních, 12 hrudních, 5 bederních, 5 křížových a 1 až 3 kostrční. Odpovídající úsek míchy, z kterého vzniká příslušný míšní nerv se nazývá míšní segment. Z horní krční míchy vystupují míšní nervy horizontálně, kolmo, dolní krční a další míšní nervy se postupně sklánějí kaudálně, až bederní, křížové a kostrční nervy vytvářejí chvost vláken (koňský chvost – cauda equina). Dochází k tomu proto, že páteř roste rychleji než mícha. Míšní nervy jsou smíšené, obsahují vlákna motorická i senzitivní.
Průřez míchou:
Na průřezu míchou je uprostřed patrný centrální kanál – canalis centralis, který je kaudálně uzavřený a kraniálně vstupuje do komorového systému mozku – IV. komory. Je vyplněn mozkomíšním mokem. Okolo centrálního kanálu je šedá hmota míšní – substantia grisea, na povrchu je bílá hmota míchy- substantia alba.
Šedá hmota míšní – substantia grisea. Má tvar motýla, který se mění v jednotlivých úsecích míchy. Rozeznáváme na ní podle polohy zadní, přední a postranní rohy míšní – cornua dorsalia (posteriores), ventralia (anteriores) et lateralia (laterales).
Zadní rohy jsou delší, štíhlejší a dosahují téměř k povrchu míchy. V jádrech zadních kořenů končí senzitivní podněty z periferie organismu (zadní kořeny míšní) a vycházejí vzestupné dráhy (axony) do vyšších center CNS a axony k buňkám předních rohů míšních.
Přední rohy jsou kratší a širší. Obsahují motorické buňky, z kterých vystupují axony (přední kořeny míšní) do svalů.
Postranní rohy jsou zřetelně vytvořeny v hrudní a křížové oblasti míchy. U krční části mají síťovitý vzhled, protože obsahují síťovité útvary – formatio reticularis medulae spinalis.
Bílá hmota míchy – substantia alba
Je na povrchu a mezi sloupci šedé hmoty. Bílá hmota míšní je rozdělena na tři párové svazky.
Zadní provazce – funiculi posteriores jsou ještě rozděleny na mediální štíhlý svazek – fasciculus gracilis (Golli) a laterální klínovitý svazek – fasciculus cuneatus (Burdachi). Zadní svazky obsahují převážně vzestupné – ascendentní dráhy.
Postranní provazce – funiculi laterales obsahují dráhy vzestupné. ascednetní i sestupné – descendentní.
Přední provazce- funiculi anteriores obsahují také dráhy jak vzestupné, tak sestupné.
Ve všech svazcích jsou kromě hlavních i vlastní svazky – fasciculi proprii posteriores, laterales et anteriores.
Struktura šedé hmoty míšní – substantia grisea
(horizontální řezy a Obrázek č. 17 Obrázek č. 18)
Šedá hmota prostupuje celou délkou míchy a je seskupena kolem centrálního kanálu do útvaru připomínajícího na příčném řezu písmeno H. V jednotlivých úsecích příčného řezu se tvar mění. Šedá hmota vybíhá dopředu jako přední sloupec (columna anterior – cornu anterius), zadní sloupec (columna posterior – zadní roh – cornu posterius), boční sloupec (columna lateralis – cornu laterale). Okolo centrálního kanálu jsou obě poloviny míchy spojeny spojkami – komisurami.
Jádra předního sloupce – columna anterior (horizontální řez 1) Obrázek č. 19 Obrázek č. 20
- 4 motorická jádra – nuclei motorii. Na jejich dendritických zónách končí neurity všech motorických systémů: dráhy pyramidové a extrapyramidové, části vestibulární dráhy a dráhy RF. Současně i některé první senzitivní neurony. Neurity motorických jader vytvářejí přední kořen – radix ventralis (motorius), který vychází v přední boční brázdě – sulcus lateralis a spolu se zadním kořenem – radix dorsalis (sensitivus) vytvářejí míšní nerv – nervus spinalis. Je jich 31–33 párů.
- malé kořenové buňky (gama – neurony), které inervují tenkým neuritem intrafusální systém nervosvalových vřetének.
- spojkové buňky, které představují interneurony vlastního míšního aparátu.
Tyto buňky tvoří somatomotorickou zónu.
Jádra zadního sloupce – columna posterior
- vlastní jádro – ncl proprius. Na buňkách tohoto jádra končí 1. senzitivní neurony, fylogeneticky staré dráhy hrubého protopatického čití (jsou to aferentní axony pseudounipolárních buňek spinálních ganglií).
Z jádra vycházejí 2. neurony přímé senzitivní dráhy. Po překřížení v přední spojce – comisura anterior se zatáčejí nahoru a končí na jádrech mezimozkového hrbolu – thalamu (dráha míchohrbolová - tr. spinothalamicus), jádrech síťové formace, FR (dráha míchosiťovaná - tractus spinoreticularis) a středomozkovém krytu – tectu (dráha míchokrytová – tractus spinotectalis)
- hrudní jádro – ncl thoracicus (jádro Stillingovo-Clarkeovo). Na buňkách tohoto jádra končí 1. senzitivní neurony hrubého protopatického čití z pohybového aparátu. Z jádra vycházejí 2. neurony nepřímé senzitivní dráhy, které se bez překřížení zatáčejí nahoru a končí na jádrech mozečku jako zadní dráha míchomozečková – tr. spinocerebellaris posterior. Jde cestou dolní mozečkové stopky – pedunculi cerebellares inferior nebo po překřížení v přední spojce – comisura anterior jako přední míchomozečková dráha – tractus spinocerebellaris anterior. Jde cestou pedunculi cerebellares anteriores.
- želatinová hmota – substantia gelatinoza (Rollandi) obsahuje buňky spojovací, které jsou interneurony nepřímých reflexů. Jejich neurity jdou vzestupně i sestupně, zkříženě i nezkříženě, končí zase v šedé hmotě a slouží ke spojení mezi segmenty
Jádra bočního sloupce – columna lateralis
- jádro prostřední boční – nucleus intermediolateralis. Patří do visceromotorické zóny. Jde o kořenové buňky, jejichž neurity jdou přes přední kořen do spinálního nervu a přes bílou spojující větev - ramus communicans albus (praeganglionaris) do ganglií truncus sympaticus. Tyto buňky jsou visceromotorické pregangliové neurony sympatické dráhy, na ně navazují neurony inervující hladké svalstvo a žlázy (viz str….)
- jádro prostřední přístřední – nukleus intermediomedialis. Jeho neurony jsou považovány za parasympatické a jejich neurity jdou k hladkému svalstvu a potním žlazám kůže, sakrální oblasti k pohlavním orgánům cestou nn. pelvici (viz str….)
Buňky obou jader tvoří visceromotorickou a viscerosenzitivní zónu.
- buňky spojovací jsou interneurony elementárního aparátu.
| Obrázky ke kapitole: Hřbetní mícha | |||
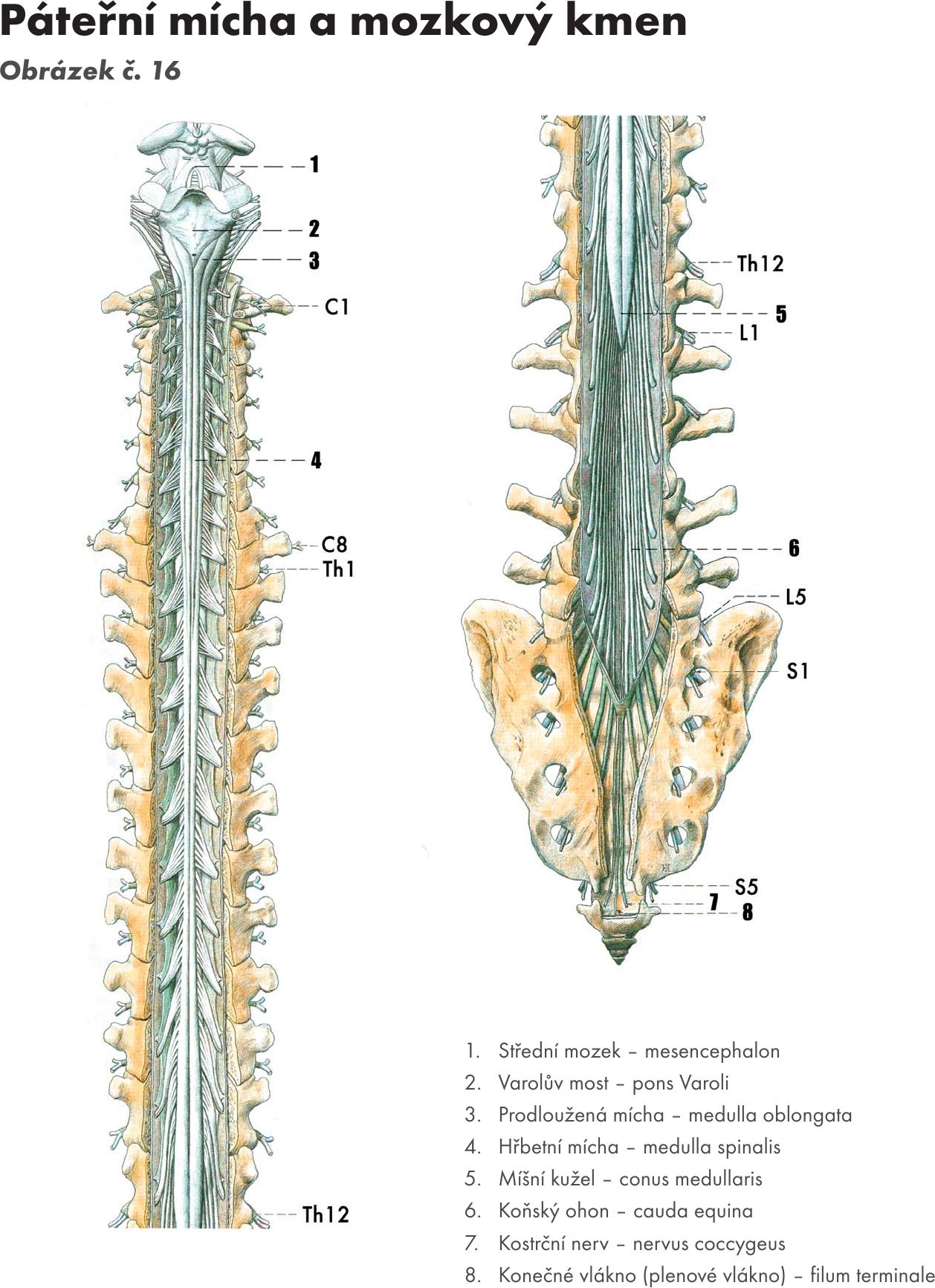
Obr. 16 Páteřní mícha a mozkový kmen
|
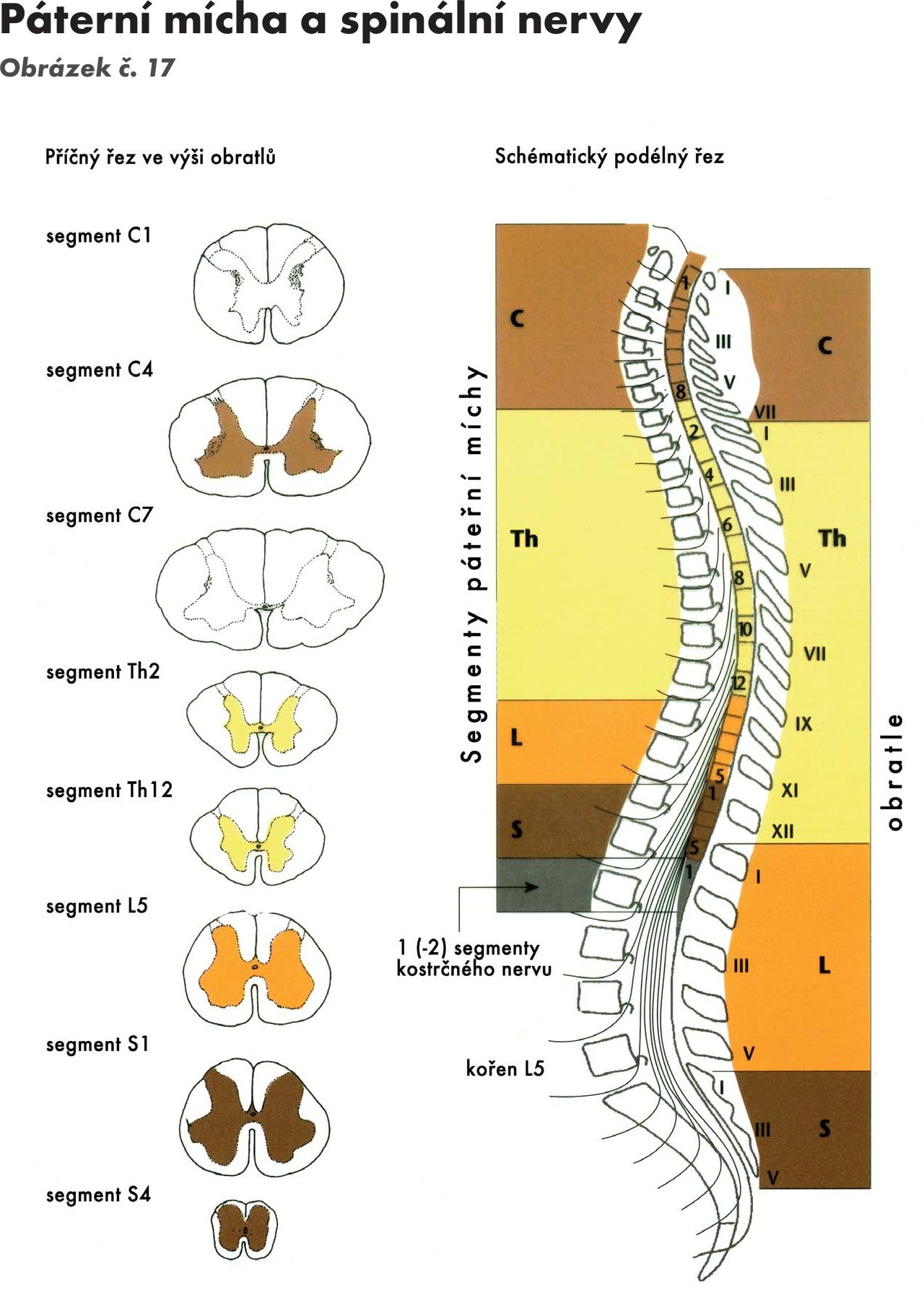
Obr. 17 Páteřní mícha a spinální nervy
|

Obr. 18 Mícha
|

Obr. 19 7 horizontálních řezů mozkového kmene
|
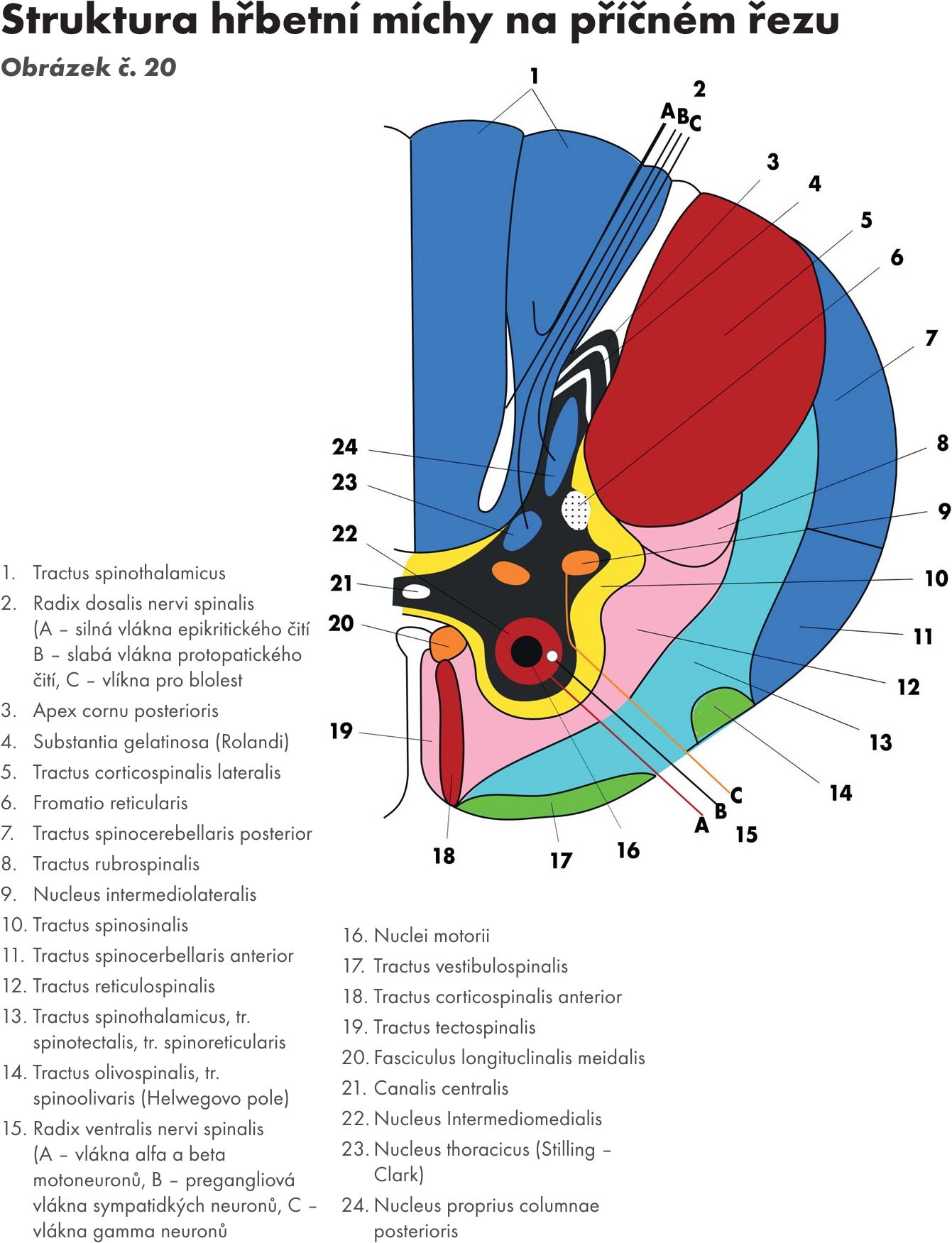
Obr. 20 Struktura hřbetní míchy na příčném řezu
|
|||
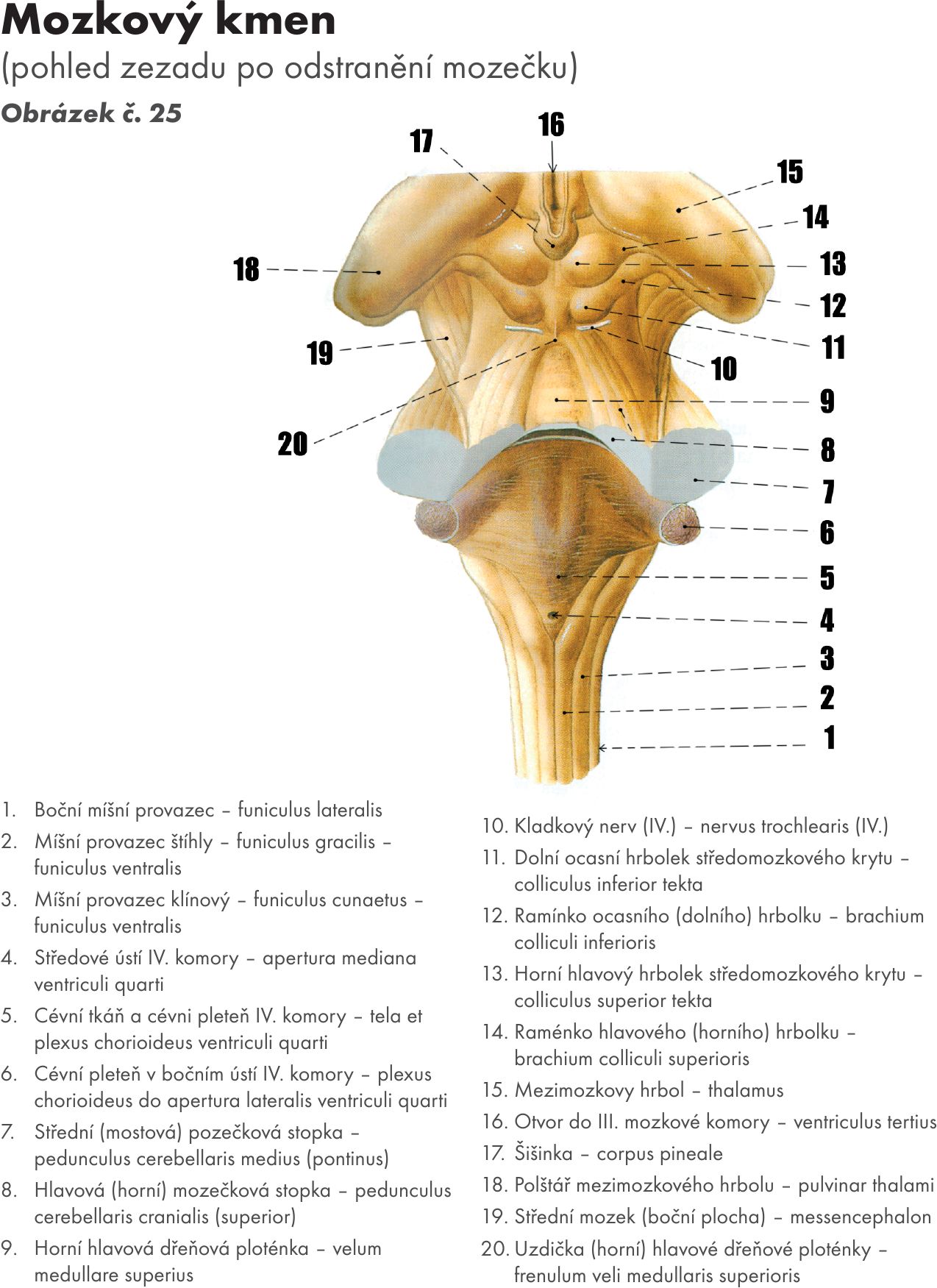
Mozkový kmen – zadní mozek
Mozkový kmen je kraniálním pokračováním míchy. Obsahuje centra nezbytně nutná pro vitální funkce. Skládá se ze tří částí: prodloužená mícha – medula oblongata, Varolův most – pons Varoli a střední mozek – mesencephalon. Dorsálně je k mozkovému kmeni připojen mozeček – cerebellum. Obrázek č. 21
Mozkový kmen s mozečkem jsou uloženy v zadní jámě lebeční. Mezi mozkovým kmenem a mozečkem je prostor, označovaný jako kosočtverečná jáma – fossa rhomboidea. Je součástí IV. komory, která je vyplněná liquorem.
Prodloužená mícha – medulla oblongata
Zevní popis
Je kraniálním pokračováním hřbetní míchy, má tvar kužele,který se rozšiřuje směrem k mostu – pontu a svou přední stranou je uložena na úbočí (svahu) – clivus ossis occipitalis.
Na ventrální ploše prodloužené míchy jsou dva symetrické protáhlé valy – pyramidy – pyramides medullae oblongatae, které jsou podmíněny průběhem pyramidové dráhy tractus corticospinalis. Hranici mezi prodlouženou míchou a hřbetní míchou tvoří křížení této dráhy pyramidové (motorické) zkřížení – decussatio pyramidum (motoria) v úrovní míšních kořenů C1.
Laterálně od pyramid leží přední boční brázda – sulcus ventrolateralis, z které vystupuje 12. hlavový nerv (N XII. – nervus hypoglossus) a zadní boční brázda – sulcu dorsolateralis, ze které vystupují nervy postranního smíšeného systému (N.IX. – glossophyrangeus, N X. – vagus a N XI. – accesorius). Mezi oběma brázdami je nápadná vyvýšenina – oliva podmíněná šedou hmotou olivárního jadra (nucleus olivaris inferior). Prodloužená mícha je od mostu oddělena brázdou – sulcu bulbopontinus, z které vystupuje 6.hlavový nerv (N VI. – nervus abducens) a laterálně v koutu mostomozečkovém odstupuje lícní a předsíňohlemýžďový nerv (N VII. – nervus facialis a N VIII. – vestibulocochlearis). Dorsální strana prodloužené míchy je pokračováním štíhlého a klínového svazku míšního (fasciculus gracilis et cuneatus) a končí štíhlým a klínovým hrbolkem (tuberculum gracilis et cuneatus). Tyto jsou podmíněny šedou hmotou stejnojmenných jader – jádro štíhlé a klínové (nucleus gracilis et cuneatus), u kterých končí vzestupné dráhy zadních provazců míšních. Laterálně od klínového hrbolku je patrný trojklanný hrbolek – tuberculum trigeminale, který je podmíněn šedou hmotou míšního jádra stejnojmenného nervu (pars spinalis nerevi trigemini – N V.). Z míchy do prodloužené míchy pokračují centrální kanál – canalis centralis, který se rozšiřuje do IV. komory – fossa rhomboidea. Rozšiření vytváří útvar podobný průřezu seříznutého husího brku upraveného na psaní – calamus scriptorius. S mozečkem je prodloužená míchy spojena dolní (ocasní) mozečkovou stopkou – peduncullus cerebellaris inferior(caudalis).
Na příčném řezu prodlouženou míchou jsou patrná jádra šedé hmoty a svazky bílé hmoty. Jádra jsou nakupením šedé hmoty (nervové buňky), bílá hmota jsou projekční nervová vlákna.
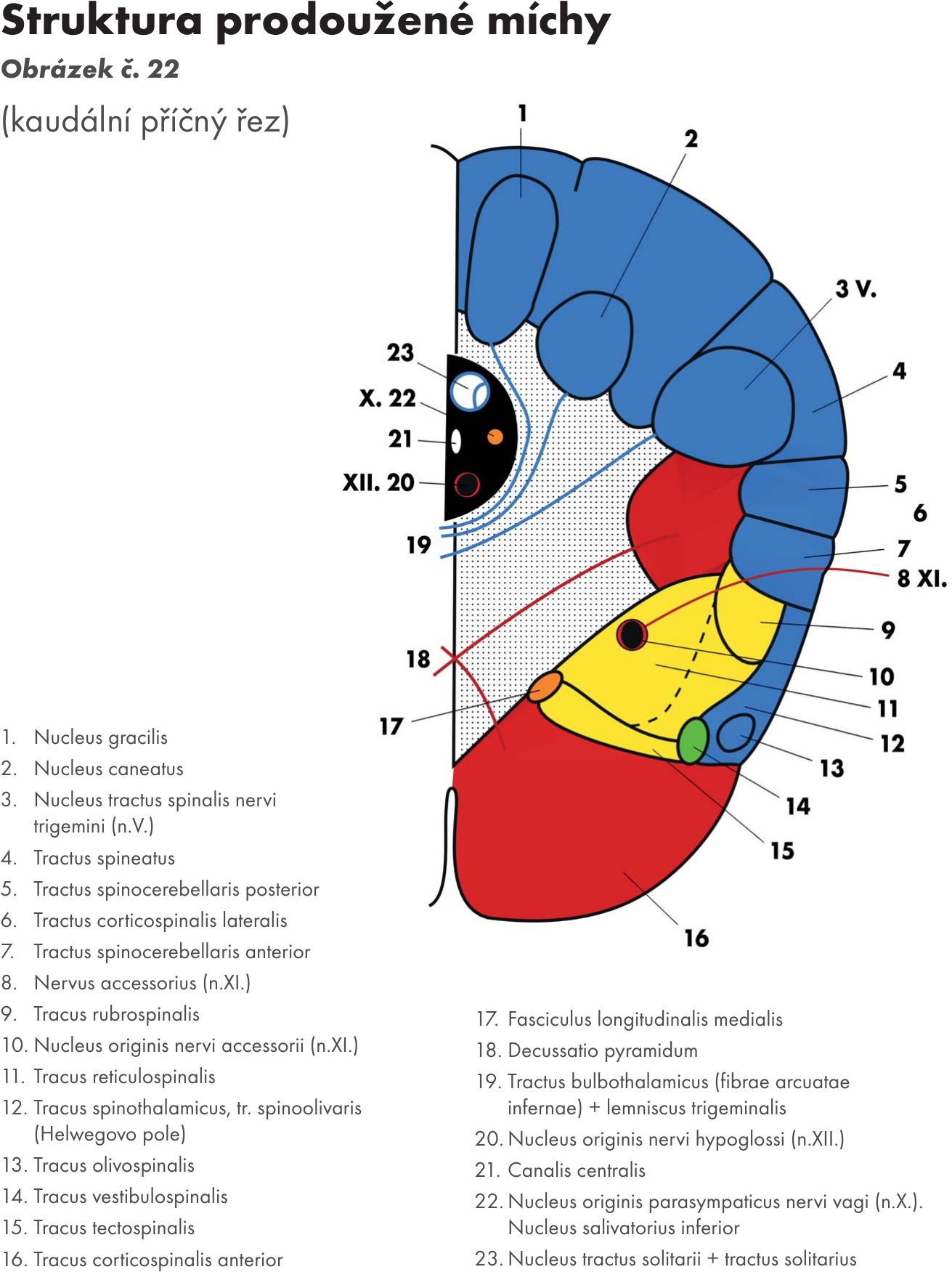
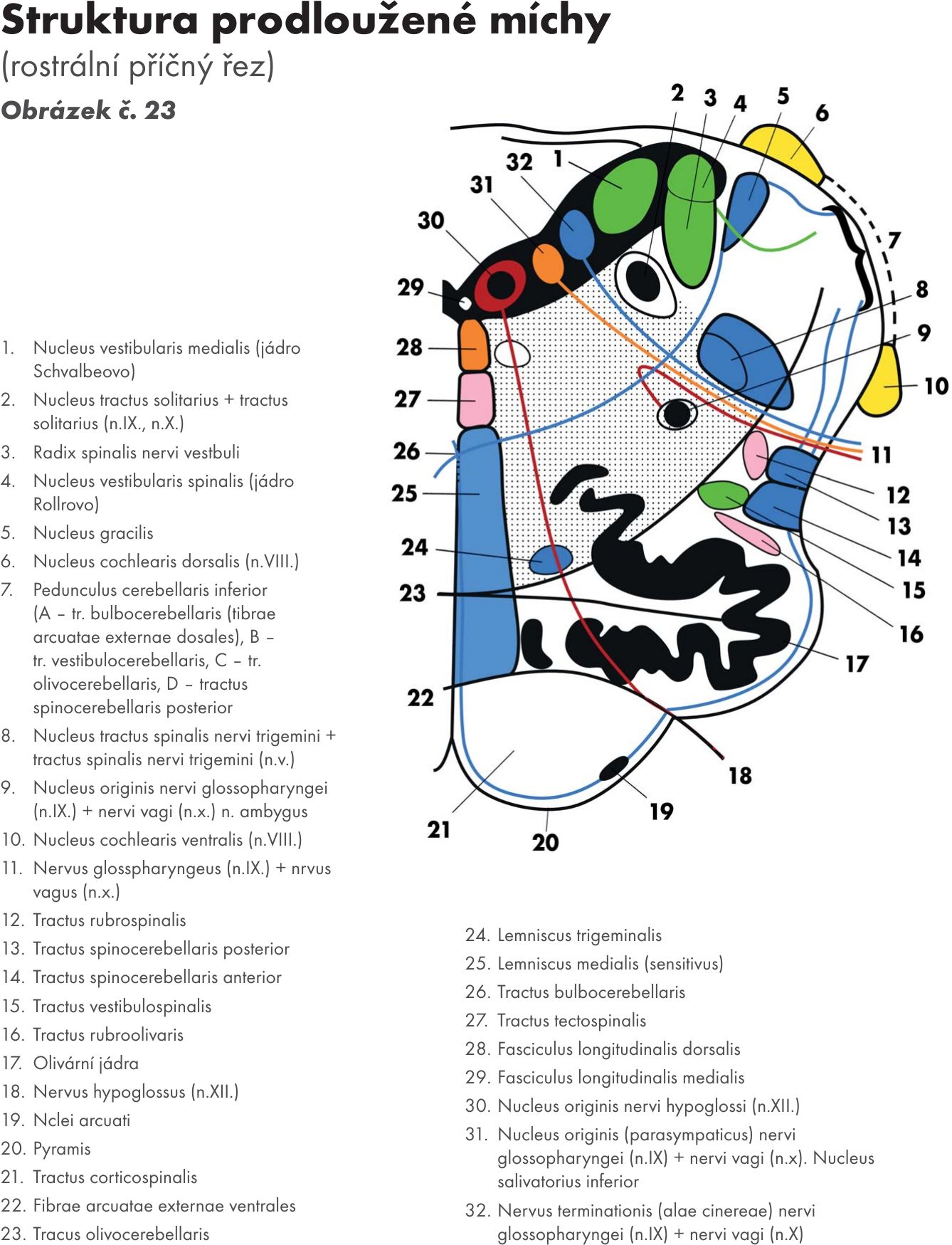
Vlastní strukturální jádra a jejich dráhy, jádra hlavových nervů (horizontální řez Obrázek č. 22 Obrázek č. 23)
Jádra zadních svazků – nuclei fasciculorum dorsalis
- jádro štíhlé – nucleus gracilis a
- jádro klínové – nucleus cuneatus.
Jádra jsou uložena v kaudální části prodloužené míchy a přepojují se na nich senzitivní dráhy zadních provazců. Jdou cestou vnitřních obloukovitých vláken – fibrae arcuatae internae, která se kříží ve střední čáře – zkřížení kliček – decussation lemniscorum. Po zkřížení vlákna stoupají mozkovým kmenem a vytvářejí prostřední kličku – lemniscus medialis. Jde o křížení druhého neuronu senzitivní dráhy (lemniskový systém) – bulvohrbolová dráha (tractus bulbothalamicus), která končí na jádrech thalamu - zadní boční přední jádro- ncl. ventralis posterolateralis. Z thalamu pak navazuje třetí neuron – dráha hrbolomozková (tractus thalamocorticalis). První neuron k jádrům prodloužené míchy přichází z míchy – dráha míchobulvová – tractus spinobulbaris.
Celou dráhu pak můžeme pojmenovat tractus spinobulbothalamocorticalis.
Přídatné klínové jádro – ncl. cuneatus accessorius. Vychází z něho bulvomozečková dráha – tractus bulbocerebellaris. Jde cestou zadních (hřbetních) vnějších obloukovitých vláken – fibrae arcuatae externae dorsales, která se někříží, a cestou předních vnějších obloukovitých vlákem – fibrae arcuatae externae ventrales, která se kříží, pokračují do mozečku dolní mozečkovou stopkou – pedunculus cerebellaris inferior.
Olivární jádra – nuclei olivares. Jsou nápadně zprohýbaná a leží na laterální straně prodloužené míchy, kde podmiňují vyklenutí – oliva.
- Přívodná vlákna (afferentní) přicházejí z míchy bočními provazci (fasciculi laterales) jako tzv. olivomíšní dráha – tractus spinoolivaris. Vede propriocepční vlákna. Další vlákna přicházejí z vyšších center CNS (z červeného jádra – ncl. ruber – dráha červenoolivární – tractus rubroolivaris, ze síťové formace – formatio reticularis - dráha síťoolivární – tractus reticuloolivaris, z mozečkových jader – tractus cerebelloolivaris).
- Odvodná vlákna (efferentní) vychází opět zkříženě jako olivomozečková dráha – tractus olivocerebellaris a jde do mozečku přes dolní mozečkovou stopku. V mozečku končí jako tzv. šplhavá vlákna na dendritech Purkyňových buněk
Jádra retikulární formace (FR). Je jich větší počet a můžeme je shrnout pod název nuclei reticulares inferiores .
Jádra hlavových nervů- nuclei nervorum cranialium pro:
nerv jazykohltanový- n. glossopharyngeus (N.XI.)
nerv bloudivý – nervus vagus (N.X.)
nerv přídatný – nervus accessorius (N. XI.)
nerv podjazykový – nervus hypoglossus (N. XII)
míšní část jádra trojklanného nervu – pars spinalis nervi trigemini (N V.)
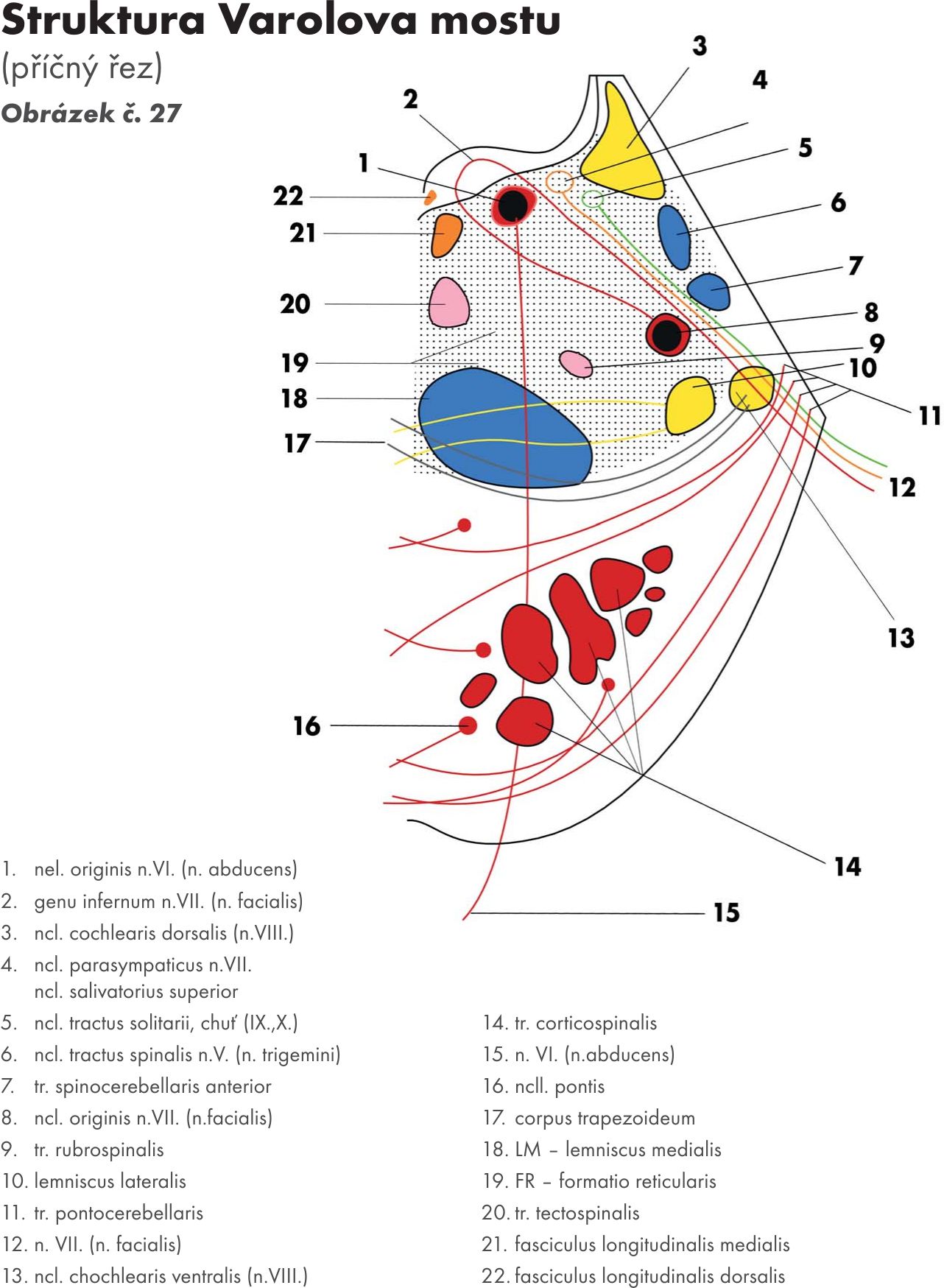
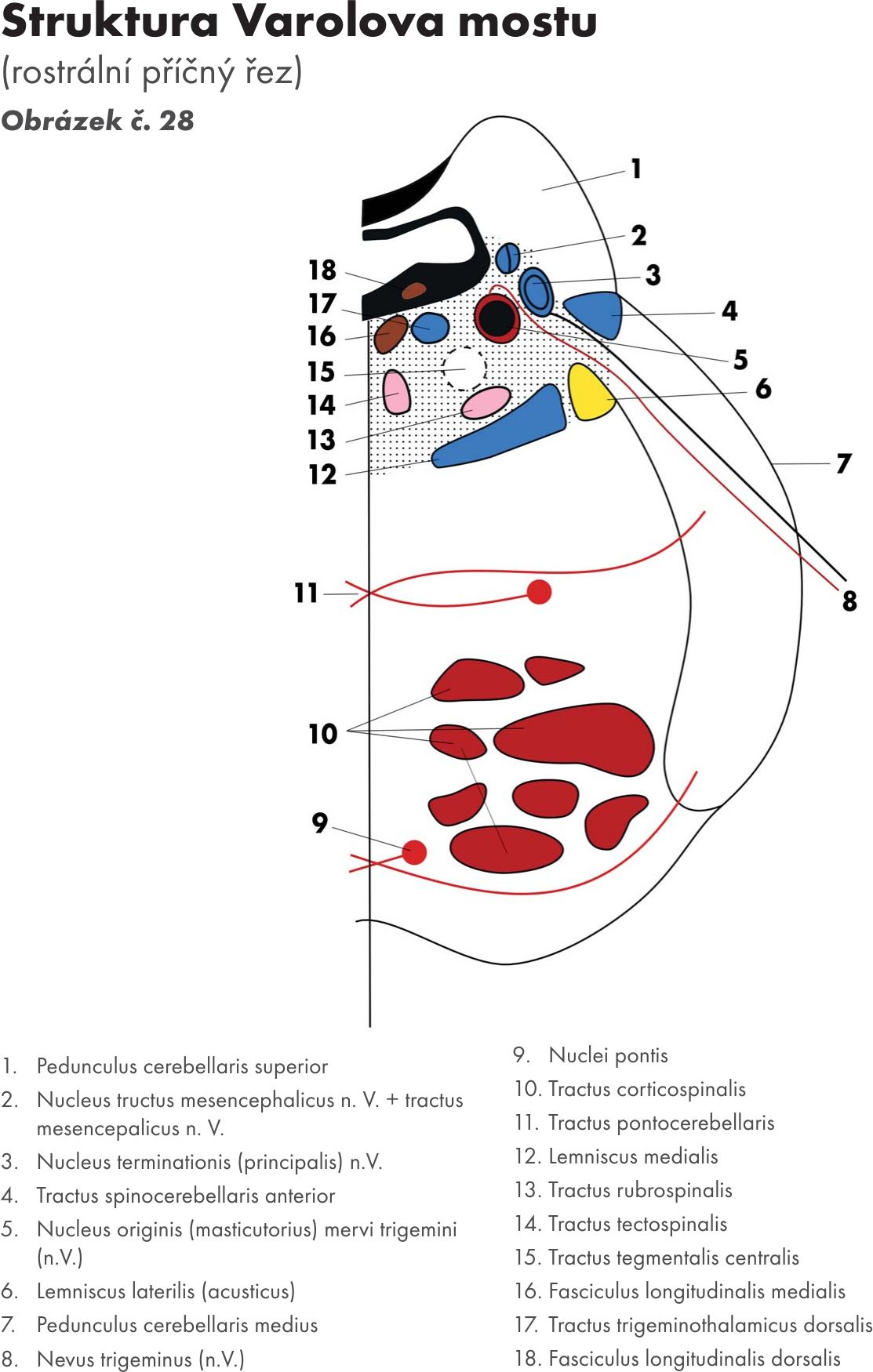
Most – Pons Varoli Obrázek č. 24
zevní popis
Je nápadný široký val na ventrální ploše mozkového kmene a je na něm patrný mělký žlábek – bazální brázda – sulcus basilaris pontis podmíněný stejnojmennou tepnou – arteria basilaris.
S mozečkem je pons spojen střední mozečkovou stopkou – pedunculus cerebellaris medius. Hranicí mezi pontem a pedunculi se uvádí spojnice výstupu obličejového nervu – n. facialis (N VII.) a trojklanného nerv – n. trigeminus (N.V), tzv. trigeminofaciální čára.. Trojklanný nerv – nervus trigeminus (N.V) vystupuje na tomto rozhraní, obličejový nerv – n. facialis v trojúhelníku (koutu) mostomozečkovém (trigonum pontocerebellare) spolu s n. vestibulocochlearis (N VIII.). V bulvomostové brázdě – sulcus bulbopontinus vystupuje n. abducens (N VI). Dorsální plochu tvoří jako u medulla oblongata spodina IV. komory – fossa rhomboidea. Obrázek č. 25
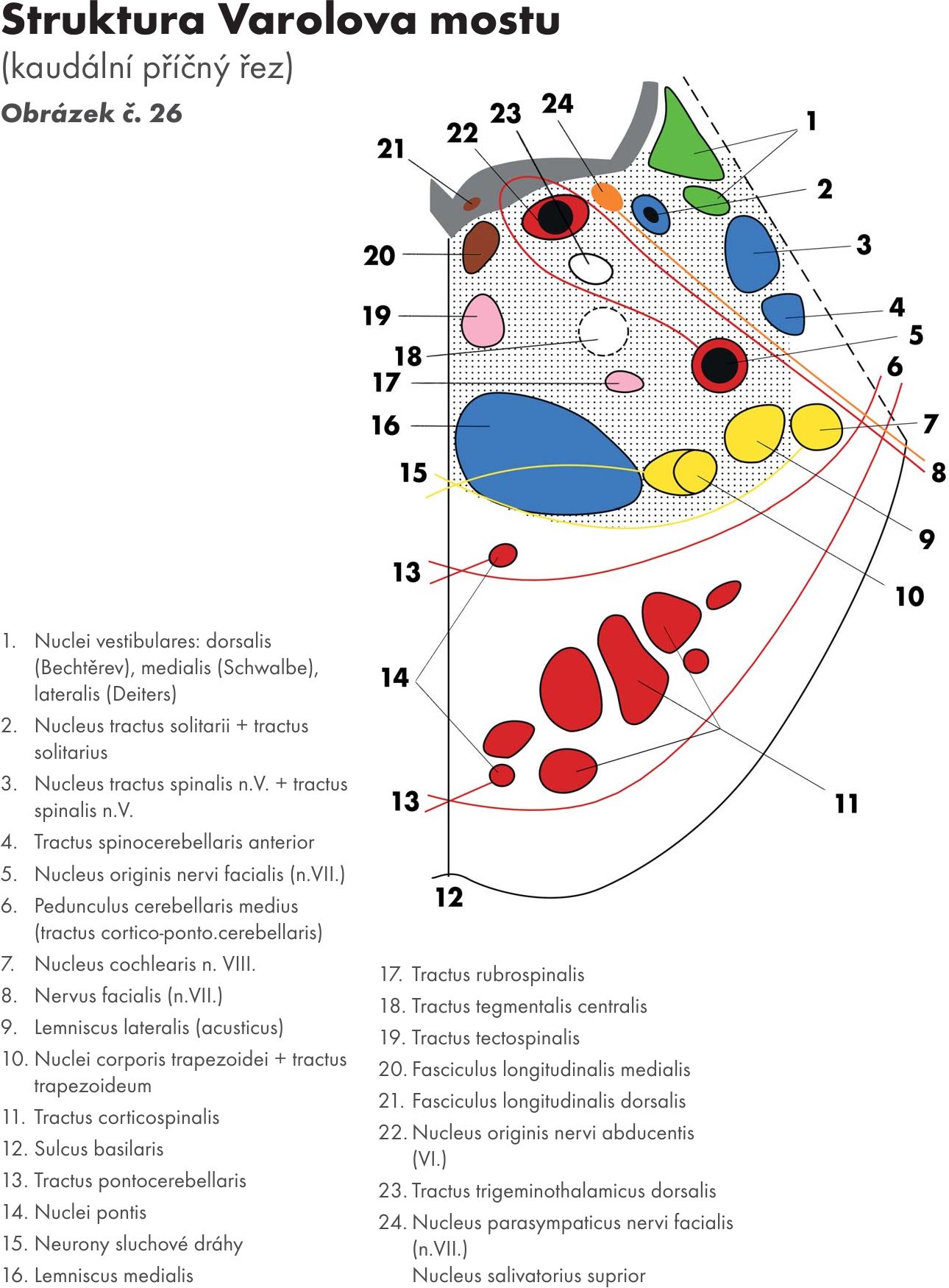
Vlastní strukturální jádra a jejich dráhy Obrázek č. 26 Obrázek č. 27 Obrázek č. 28
Mostová jádra – nuclei pontis, leží v basální části Varolova mostu. Prostupuje jimi koromíšní dráha- tractus corticospinalis, která ji rozděluje na větší počet svazečků – dlouhá mostní dráha – fibrae pontis longitudinalis – svazky roztříštěných pyramid.
- Přívodná vlákna (afferentní) koromostní vlákna (fibrae corticopontinae – tractus corticopontinus) Přicházejí z téměř celé kůry mozkové.
- Odvodná vlákna (efferentní) koromostní vlákna po zkřížení pokračují do mozečku jako vlákna mostomozečková (fibrae pontocerebellares – tractus pontocerebellaris), kde končí opět na dendritech Purkyňových buněk. Procházejí do mozečku skrze střední mozečkovou stopku, jejíž jsou hlavní součástí. Některá vlákna mostomozečková se přepojují na:
Obloukovitá jádra – nuclei arcuati, která leží pod povrchem pyramid. V předních vnějších obloukovitých vláknech – fibrae arcuatae externae ventrales. Po zkřížení vstupují do mozečku přes dolní mozečkovou stopku pedunculus cerebellaris inferior.
Jádra retikulární formace (FR) – nuclei reticulares medii
Jádra hlavových nervů – nuclei nervorum cranialium
pro:
nerv trojklanný – nervus trigeminus (N V)
nerv odtahující – nervus abducens (N VI)
nerv lícní – nervus facialis (N VII)
nerv sluchově rovnovážný – nervus statoacusticus (N VIII.) čili nerv rovnovážný a sluchový -. nervus vestibulocochlearis.
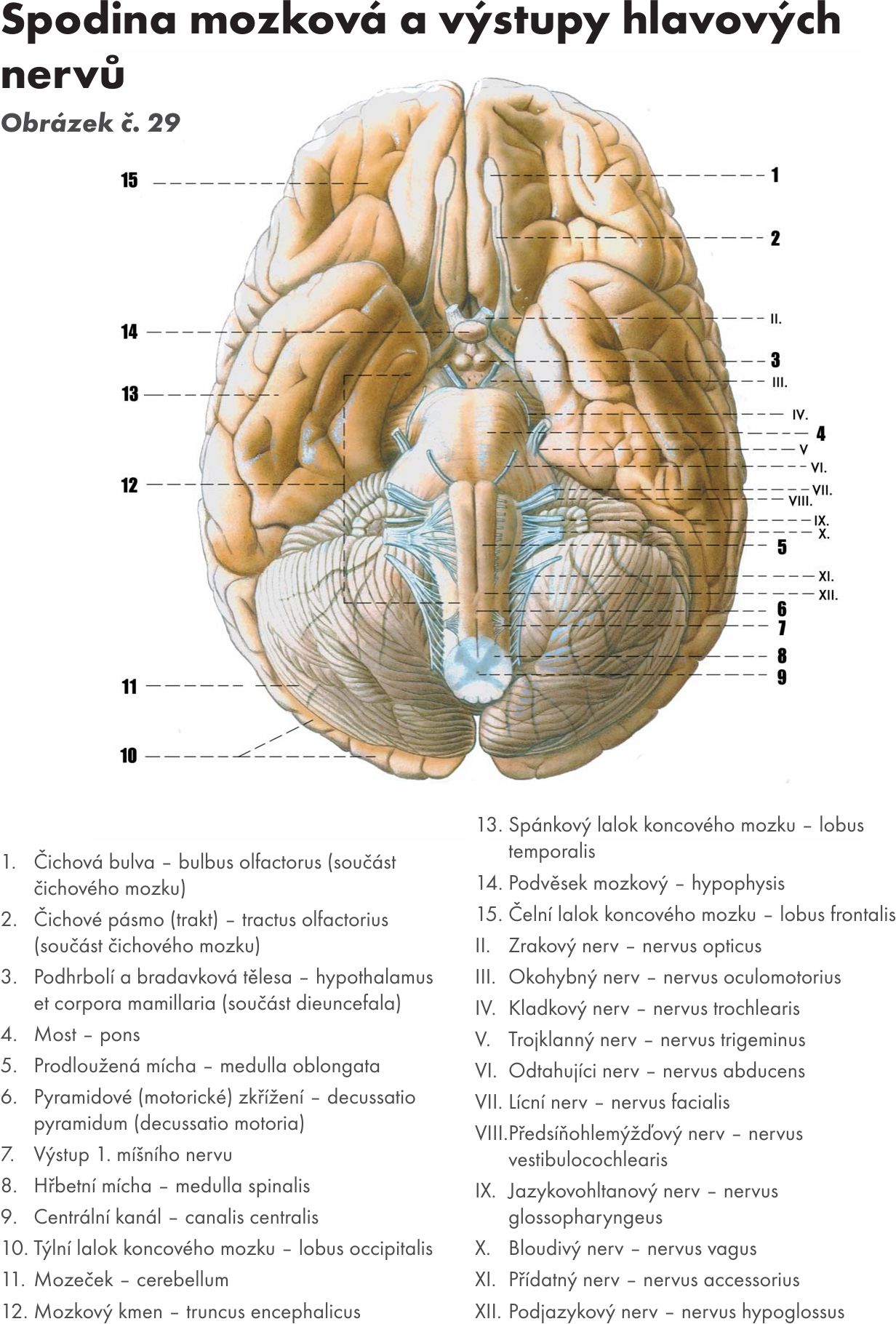
Jejich výstup ze spodiny mozkové Obrázek č. 29
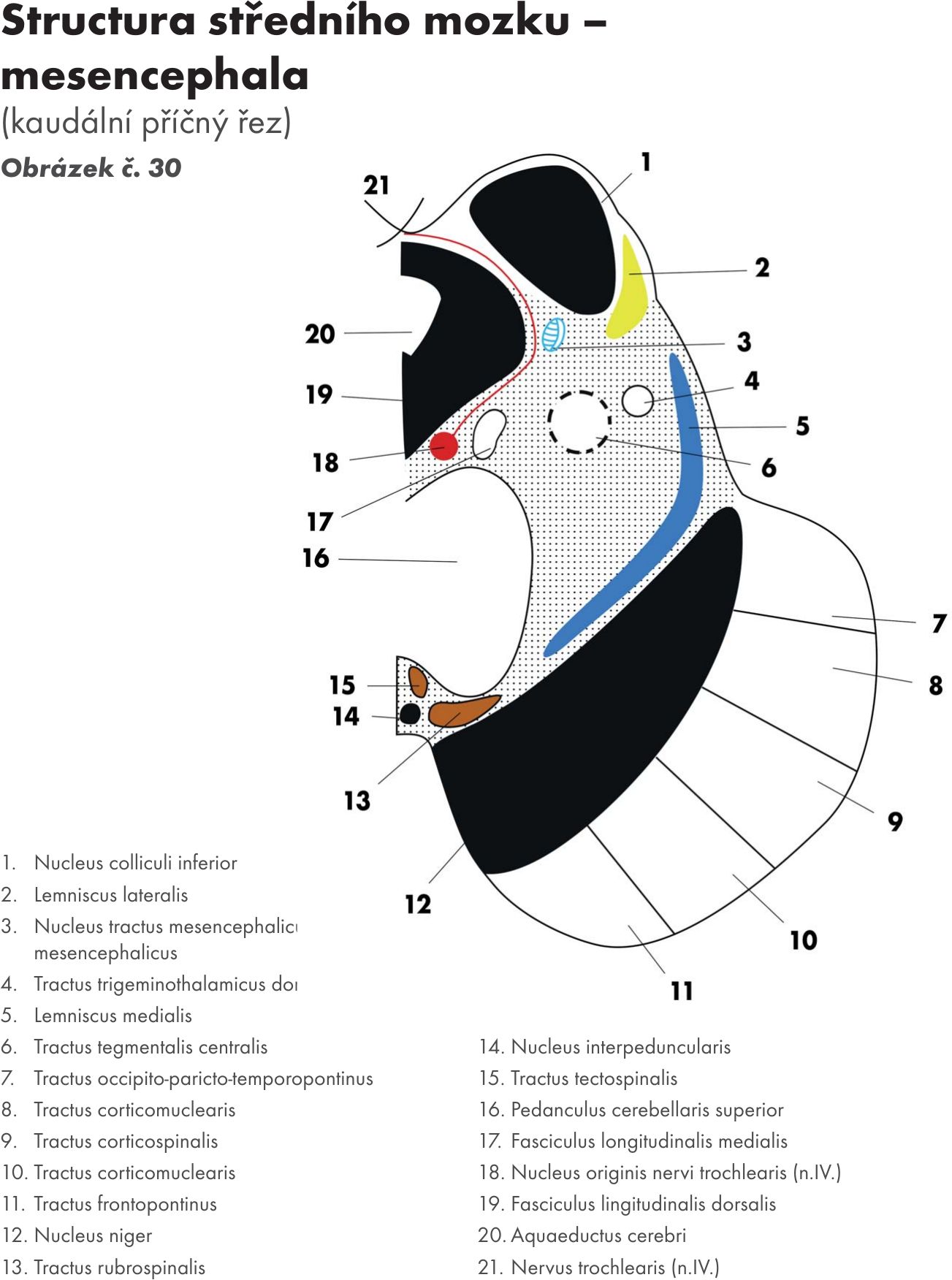
Střední mozek – mesencephalon
zevní popis
Základní části mesencephala jsou tři zřetelně rozlišené etáže:
- ramena mozku – crura cerebri ventrálně uložená, nad nimi se rozkládá
- středomozková pokrývka (čepec) – tegmentum mesencephali, nad kterým leží
- středomozkové krytí – čtverohrbolí – lamina tecti – tectum mesencephali, na kterém leží dva páry hrbolků – horní hrbolky – colliculi superiores a dolní hrbolky- colliculis inferiores, které jsou podmíněny stejnojmennými jádry.
Horní hrbolky jsou zapojeny do zrakové dráhy, dolní do dráhy sluchové. Oba páry hrbolků jsou spojené s mezimozkem oblými párovými raménky – brachia colliculi superiores et inferiores, která probíhají laterálně a končí v prostředních a bočních kolénkových tělíscích – corpus geniculatum mediale et laterale thalami.
Na řezu středním mozkem je mezi tectem a tegmentem viditelný kanálek, mokovod – aqueductus mesencephali Sylvii, který spojuje III. a IV.komoru mozkovou, které jsou vyplněné mozkomíšním mokem.
S mozečkem je mesecephalon spojen horními mozečkovými stopkami- pedunculi cerebellares superiores, mezi kterými je napjatá ploténka bílé hmoty – velum medulare superiores, která tvoří část stropu IV. komory mozkové- tegmen ventriculi quarti.
Tak jako prodloužená mícha a most tvoří i sřední mozek spodinu IV. komory. Ze středního mozku vystupují dva hlavové nervy :
- okohybný nerv – n. oculomotorius (N III.), vystupuje v mezistopkové jámě a
- kladkový nerv – n. trochlearis (N IV.), je to jediný hlavový nerv, který vystupuje na dorsální straně mozkového kmene.
Vlastní strukturální jádra a jejich dráhy
- Středomozkový kryt – tectum: Obrázek č. 30 Obrázek č. 31
- Jádra horních hrbolků – ncl. colliculi superiores. Mají žíhaný vzhled, střídají se v nich vrstvičky bílé a šedé hmoty. Jsou součásti zrakové dráhy a přijímají (afferentní dráhy): dráha míchokrytová – tractus spinotectalis a vlákna z uzdičky jader – ncl. habenulae – uzdičkokrytová dráha – tr.habenulotectalis. V tomto podkorovém zrakovém centru se zrakové signály integrují se signály z dalších oblasti CNS.
- Pretektální jádra – ncll. praetectales. Leží těsně před colliculis superiores v přetektální oblasti. Tato jádra dostávají odbočky ze zrakové drány a jsou přepojovací stanicí pupilárního reflexu.
- Jádra dolních hrbolků – ncl. colliculi inferiores, vytvářejí po jednom větším jádru, jsou přepojovací stanicí sluchové dráhy ze zevní kličky – lemniscus lateralis.
Efferentní dráhy :
- Krytomíšní – tr.tectospinalis, končí na motorických jádrech míchy (viz str., jejich vlákna podporuje tr. tectoreticularis
- Krytobulvární (bulva, cibule) – tr. tectobulbaris, končí na motorických jádrech hlavových nervů.
- Krytosíťovaná dráha – tr. tectoreticularis, končí na jádrech FR (formatio reticularis) středního mozku. Převádí zrakové impulsy na jádra FR a navazuje na ni tr. reticulospinalis.
- Krytomostní dráha – tr. tectopontinus, končí na laterálních jádrech FR v pontu.
- Středomozková pokryvka- tegmentum
Červené jádro – nucleus ruber. Je oválné jádro ležící mediálně. Má načervenalou barvu, kterou působí pigment s obsahem železa. Většinu objemu jádra tvoří u člověka a primátů rostrální malobuněčná část – pars parvocellularis. Kaudální velkobuněčná část – pars magnocellularis převažuje u ostatních savců.
- Afferetní dráhy:
- mozečkočervená – tr. cerebellorubralis, přichází z mozečku přes horní mozečkovou stopku
- koročervená – tr. corticorubralis, přichází z kůry mozkové
- bledočervená dráha – tr. pallidorubralis, přichází z basálních ganglií.
- Efferentní dráhy :
- červenomíšní – tr. rubrospinalis
- červenolůžková – tr. rubrothalamicus
- červenokorová dráha- tr. rubrocorticalis
- červenomozečková dráha- tr. rubrocerebellais, která jde do mozečku přes horní mozečkovou stopku.
Černá hmota – substatia nigra. Je protáhlé jádro černé barvy. Leží na rozhraní mezi tegmentum mesencephali a ramena mozku – crura cerebri.
Dělí se na: dorsálnější hutnou část – pars compacta a
ventrální síťovanou část – pars reticularis.
Černou barvu hutné části - pars compacta tvoří shluky tmavě pigmentovaných neuronů, které produkují radiátor dopamin, který je axonálním prouděním transportován do basálních ganglií (dopaminergní systém mozku). Vedlejším, produktem při syntéze dopaminu je černý pigment – neuromelanin.
Ventrální síťovanou část – pars reticularis, která je tvořena řidčeji rozloženými neurony, které zasahují mezi drány v crura cerebri. Buňky obsahují žlutohnědý pigment (lipofuscin)
- Afferentní dráhy:
- dráha bledočerná - tr. pallidonigralis
- drána žíhanočerná – tr. strionigralis. Končí na pars reticularis. Vlákna mají inhibiční vliv na neurony pallida a na neurony nigrální.
- Efferentní vlákna:
- drána černožíhaná – tr. nigrostriatus. Vlákna vystupují z pars compacta a transportují dopamin, který je nutný k normální funkci striata. Při jeho sníženém přísunu do striata se objevují příznaky parkinsonismu.
- dráha černosíťovaná – tr. nigroreticularis. Vlákna vystupují z pars reticularis a jsou z funkčního hlediska inhibiční.
- dráha černolůžková – tr. nigrothalamicus.
Mezistopkové jádro – nucleus interpeduncularis je malé párové jádro uložené pod spodinou mezistopkové jámy – fossa interpeduncularis. Jádra v něm jsou zapojena do tzv. limbického systému. Aferetní vlákna přicházejí z uzdičkomezistopkové dráhy – tr. habenulointerpeduncularis, eferentní vlákna míří do retikulární formace (FR).
Centrální šedá hmota- substatia grisea centralis je šedá hmota obkružující středomozkový mokovod – aqueaductus Sylvii. Kraniálně zasahuje do mezimozku ke stěně III. komory, kaudálně až pod spodinu IV. komory. Funkčně připomíná retikulární formaci, má bohaté afferentní a efferentní spoje. Svými sestupnými spoji ovlivňuje přenos signálů bolesti v dráze míšně siťovanolůžkové – tractus spinoreticulothalamicus.
K centrální šedé hmotě se v rostrální oblasti středního mozku přikládají dvě jádra vmezeřená Cajalovo jádro – ncl. interstitialis (Cajali) a Darkševičovo jádro – ncl. Darschewitschi.
Affferetní vlákna přicházejí z přístředního podélného svazku – fasciculus longitudinalis medialis (vlákna z vestibulárních jader).
Efferetní vlákna z obou jader jdou k vestibulárním jádrům,k FR a k jádrům okohybných hlavových nervů (N III,IV,VI.).
Jádra retikulární formace (FR) – nuclei reticulares superiores.
Jádra hlavových nervů – nuclei nervorum cranialium pro nerv okohybný – n oculomotorius (N III) a nerv kladkový – n. trochlearis (N IV)
- Afferetní dráhy:
- Ramena mozku – crura cerebri.
Párová ramena uložená na spodině mesencephala. Jsou tvořena bílou hmotou, která obsahuje sestupné dráhy:
- Kortikomíšní dráha – tractus corticospinalis jsou vlákna pyramidových drah. Prochází středem crura cerebri mesencephala a spojují kůru mozkovou s motorickými míšními jádry
- Korojádrová dráha – tractus corticonuclearis sestupuje po mediální i laterální straně tr. corticospinalis a spojuje kůru mozkovou s motorickými jádry – ncl. originis hlavových nervů.
- Koromostová drána – tractus corticopontinus. Jednak vláka procházejí skrze crura cerebri nejmediálněji – fibrae pontinae (Arnoldova dráha), druhá vlákna procházejí nejlaterálněji fibrae parietooccipitopontinae (Türckova dráha). Vlákna vedou z drah mozkové kůry do nuclei pontis, odkud navazuje jako dráha mostomozečková – tr. pontocerebellaris. Jediná dráha, která vede do mozečku přes pedunculus cerebellaris medius.
Kromě již jmenovaných drah prochází kmenem mozkovým typické okrsky bílé hmoty. Jsou tvořeny myelinizovanými i nemyelinizovanými vlákny. Tvoří tyto dráhy mozkového kmene:
- Přístřední klička- lemniscus medialis: vede vzestupné dráhy senzitivní povahy z ncl gracilis et cuneatus z prodloužení míchy – medula oblongata a z ncl. proprius z hřbetní míchy- medula spinalis a ze senzitivních jader hlavových nervů. Největší je trojklanná klička – lemniscus trigeminalis. Na řezu středním mozkem leží svazek dorsolaterálně od nucleus ruber. Dráhy v něm probíhající končí v thalamu – lůžku.
- Zevní klička – lemniscus lateralis. Její vlákna jsou pokračováním sluchové dráhy z kochleárních jader a jsou přerušena předním a zadním trapézovitým (stolečku podobným) jádrem a končí v collliculi inferiores. Leží dorsolaterálně od lemniscus medialis.
- Prostřední podélný svazek – fasciculus longitudinalis medialis. Leží ventrálně od substantia grisea centralis. Obsahuje vzestupná i sestupná vlákna a začíná v rostrálním mesencephalu (v horní části středního mozku). Propojuje Darkševičovo a Cajalovo jádro, jádra vestibulární a jádra okohybných nervů.
- Hřbetní podélný svazek – fascisulus longitudinalis dorsalis. Vystupuje z hypothalamu – podlůžka. Probíhá tegmentem dorsálně od fasciculus longitudinalis medialis těsně pod spodinou IV. mozkové komory. Většina vláken končí v parasympatických jádrech hlavových nervů (N III,VII,IX,X).
- Centrální pokrývkový svazek – fasciculus centralis tegmenti. Vystupuje párově z mesencephala z malobuněčné části červeného jádra – ncl. ruber, leží při jeho dorsální straně. Axony tohoto objemného svazku bílé hmoty sestupují v dorsálním olivárním jádru – nucleus olivaris inferior, v přídatném klínovém jádru – nucleus cuneuatus accesorius a v jádrech retikulární formace (FR).
- Dolní, střední a horní mozečkové stopky – pedunculi cerebellares inferiores, medii et superiores. Tyto stopky spojují jednotlivé části kmene mozkového s mozečkem a dráhy aferetní i eferetní jimi procházející budou uvedeny u mozečku – cerebellum.
- Středomozkový kryt – tectum: Obrázek č. 30 Obrázek č. 31
Jádra hlavových nervů – nuclei nervorum cranialium pro
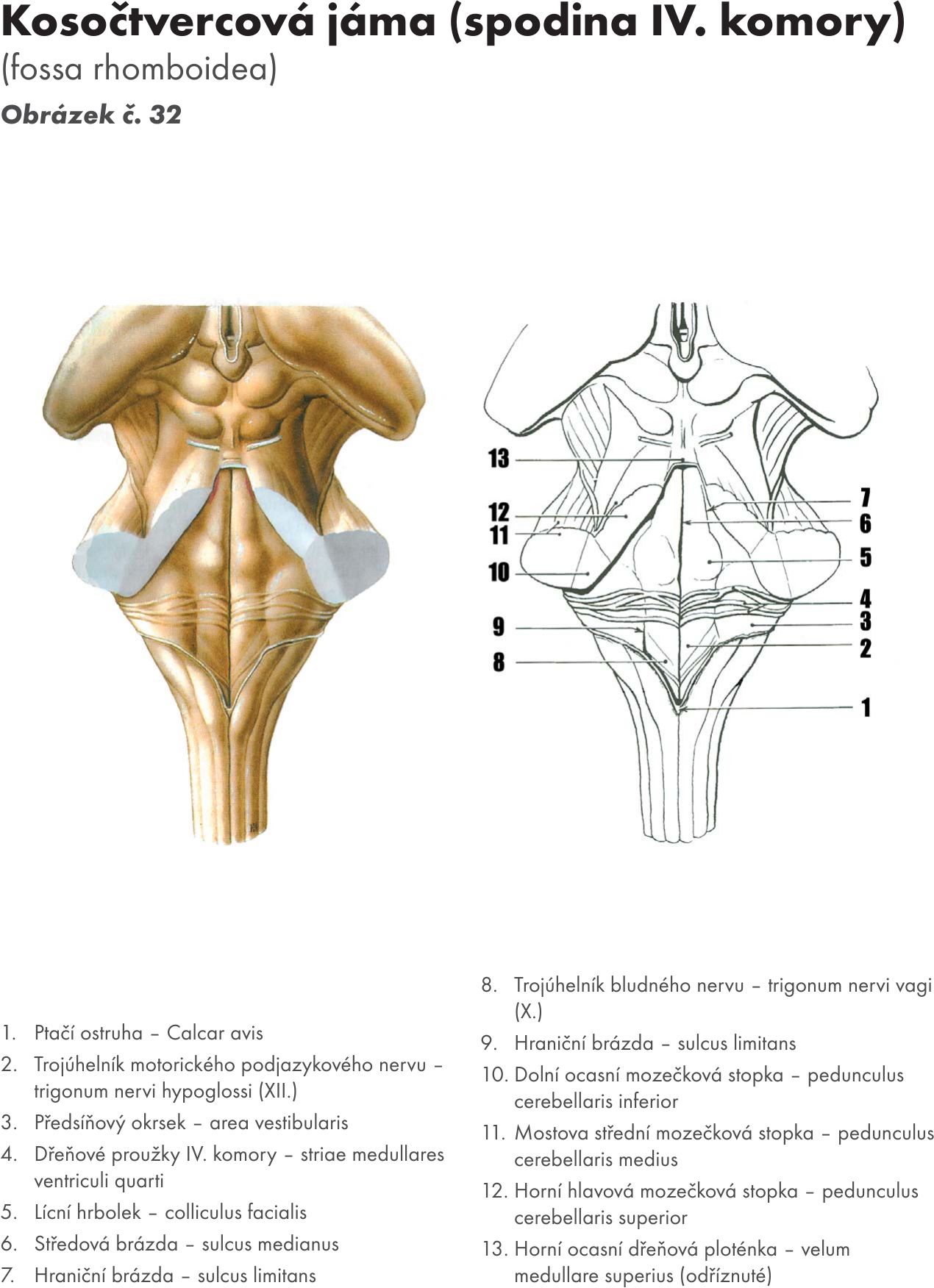
nerv okohybný, n. oculopeotorius (N III.) a nerv kladkový, n. trochleris (N. IV.)Kosočtverečná jáma – fossa rhomboidea Obrázek č. 32
- zevní popis
Tvoří spodinu IV. komory mozkové – ventriculus quadratus. Přehlédnout ji můžeme pouze po odříznutí mozečku – cerebellum. Zašíná v canalis centralis míchy, rozšiřuje se pod mozečkem a končí v mokovodu středního mozku – aquaeductus mesencephali (Sylvii). Je tvořena dorsální plochou prodloužení míchy, Varlova mostu a středního mozku. Ve střední části se rozšiřuje v boční vychlípení – recessus lateralis – nejširší místo komory. Kaudálně je ohraničena dolními (ocasními) mozečkovými stopkami – pudunculus cerebellares inferiores, uprostřed středními mozečkovými stopkami – pedunculus cerebellares medii a vpředu horními mozečkovými stopkami – pedunculus cerebellares superiores. Středovou brázdou – sulcus medianus je rozdělena na dvě poloviny. Po stranách brázdy jsou patrné prostřední vyvýšeniny, valy – eminentia mediales, které laterálně ohraničují hraniční brázdy- sulci limitantes. Během vývoje oddělují bazální a alární ploténku. Po rozevření neurální strubice leží bazální ploténka (motorická) mediálně od sulcu limitans, alární ploténka (senzitivní) laterálně od sulcus limitans.
Na spodině IV.komory jsou patrny další významné útvary :
- V kaudální části fossa rhomboidea nad medula oblongata je podjazykový trojúhelník – trigonum nervi hypoglossi. Pod ním je motorické jádro n. hypoglossus (N XII.).
- Bludný troúhelník – trigonum nervi vagi. Pod ním je dorsální jádro n. vagus (N X.)
- Ve střední části fossa rhomboidea nad pontem je
lícní hrbolek – colliculus facialis. Leží mediálně od sulcus limitans. Je podmíněn kolínkovým ohbím vláken n. facialis (N VII.) kolem jádra n. abducens (N VI.).
- Dřeňové proužky – striae medullares. Jsou to jemné světlé proužky viditelné pod colliculus facialis. Obtáčejí pedunculus cerebellares superiores, zanořují se do střední brázdy - sulcus medianus. Obsahují části vláken sluchové dráhy z nucleus cochlearis dorsalis
- Předsíňový okrsek – area vestibularis je vyvýšenina ležící v recessus lateralis ventriculi IV. (quarti) a leží pod ní jádra n. vestibulocochlearis (N VIII)
- V rostální části fossa rhomboidea leží namodralé políčko – locus coeruleus. Leží pod ním stejnojmenné jádro, které obsahuje noradrenergní, tmavě pigmentované buňky.
Pod povrchem spodiny IV. komory (fossa rhomboidea) jsou uložena jádra hlavových nervů.
Jádra hlavových nervů – nuclei nervorum cranialium
Jsou uspořádána do dvou řad. Tyto longitudinální pruhy jader odpovídají základnímu vývojovému členění nervové trubice, to je ploténka basální – motorická zóna a ploténka alární – senzitivní zóna.
Mediální řada jader je mezi sulcu medianus a hraniční brázdou – sulcus limitans a odpovídají basální ploténce – jádra motorická:
- somatomotorická – inervují svaly, které pocházejí ze somitů
- branchiomotorická – inervují svaly, které pocházejí vývojově ze žaberních oblouků
- visceromotorická – parasympatická.
Laterálně od sulcus limitans je ploténka alární. Zde leží jádra senzitivní:
- somatosenzitivní – přijímají informace z oblasti samotické
- viscerosenzitivní – přijímají informace z oblasti vnitřních orgánů
- senzorická – z čiv.
Motorická jádra nervů se označují jako počáteční jádro (nucleus originis) nebo motorická jádra (nucleus motorius). Jádra odpovídají skupinám motoneuronů v předních a v postranních sloupcích míšních. Jejich axony tvoří motorická vlákna příslušného nervu a končí na neuromotorických ploténkách svalových vláken.
Senzitivní jádra nervů se označují jako zakončující jádra (nucleus terminationis) příslušného nervu a odpovídají jádrům v zadních sloupcích míšních. Jejich axony pokračují jako ascendentní (vzestupné) dráhy k vyšším centrům CNS.
Prodloužená mícha – medula oblongata:
Somatomotorická zóna:
- Nucleus originis nervi hypoglossi (NXII) – jádro nervu podjazykového. Leží pod podjazykovým trojúhelníkem – trigonum nervi hypoglossi, je motorické jádro inervující svaly jazyka.
Branchiomotorická zóna:
- Nucleus ambiguus (dvojaké jádro). Leží pod bludným trojúhelníkem – trigonum nervi vagi. Je to motorické jádro pro nervy IX,X,XI. Inervují svaly: N IX. (měkkého patra, mimo m.tensor veli palatini, hltanové branky, úžiny hltanové – isthmus faucium a svaly hltanu), N X. (svaly hltanu a hrtanu) a N XI. (m. sternocleidomastoideus – kývač hlavy a m. trapezius – sval kápový).
Visceromotorická zóna:
- Dolní slinné jádro – ncl. salivatorius inferior pro N IX,N X.
Viscerosenzitivní zóna:
- Jádro popelavé – ncl. cinereus pro N XI,N X.
Somatosenzitivní zóna:
- Míšní jádro trojklanného nervu – ncl. spinalis nervi trigemini (radix spinalis N V.). Končí v něm vlákna vedoucí signály bolesti, tepla, chladu a propriocepce ze somatosenzitivních oblastí N V. Končí v něm somatosenzitivní vlákna nejen z ganglion trigeminale, ale i ganglií N VII,NIX., N X. K trigeminovému jádru odbočují až v mozkovém kmeni.
Sensorická zóna:
- Osamocené jádr o- ncl. solitarius. Je protáhlé jádro uložené zevně při sulcus limitans. Kraniální úsek tohoto jádra se nazývá jádro chuťové – ncl. gustatorius a přijímá chuťová vlákna z ganglí N VII., IX.,N X..
Most – pons Varoli:
Somatomotorická zóna:
- Jádro odtahujícího nervu – nucleus originis nervi abducentis. Je vloženo pod lícním hrbolkem – colliculus facialis, který je podmíněn kolínkovitým ohbím vláken lícního nervu – nervus facialis (N.VII), která jádro N VI. obtáčí. Přijímá korojádrová vlákna – fibrae coritconucleares. Z jádra vystupují axony cestou N VI pro přímý zevní sval oční – m. rectus bulbi lateralis
Branchiomotorická zóna:
- Motorické jádro trojklanného nervu – ncl. motorius (originis) n. trigemini (NV.), jádro žvýkací – ncl. masticatorius. Efferentní vlákna motoricky inervují žvýkací svaly, m. myohyoideus, m. tensor veli palatini a m. tensor tympani.
- Motorické jádro lícního nervu – ncl. motorius n. facialis (N VII). Motorická vlákna jsou určena pro svalstvo II. žeberního oblouku – svyl mimické.
Visceromotorická zóna:
- Horní slinné jádro – ncl. salivatorius superior pro N VII
Viscerosenzitivní zóna:
- Senzitivní vlákna N VII odvádějí senzitivní vjemy ze sliznice středoušní dutiny.
Somatosenzitivní zóna:
- Mostní jádro trojklanného nervu – ncl. pontinus (principalis) N V. Končí v něm typická somatosenzitivní vlákna z trojklanné nervové uzliny – ganglion trigeminale (Gasseri). Přijímá podněty citlivosti, hmatu, vibrace a propriocenpce ze somatosenzitivní oblasti N V.
- Somatosenzitivní vlákna N VII pro inervaci kůže boltce. Vlákna míří do míšního jádra ncl. spinalis N V.
Sensorická zóna:
- Chuťové jádro – ncl. gustatorius. Je kraniální úsek osamoceného jádra- ncl. solitarius, které leží v prodloužení míše. Přijímá chuťová vlákna z ganglií N VII,IX,X..
- Předsíňová jádra – ncll.vestibulares. Leží laterálně na spodině IV. komory pod předsíňovým okrskem – area vestibularis. Přijímají podněty z vnitřního ucha, z orgánů rovnováhy (N. VIII) a informují o poloze hlavy a jejich změnách (sklonění) a o otáčivých pohybech hlavy (zrychlení pohybu):
- přístřední předsíňové jádro – ncl. vestibularis medialis (Schwalbeovo jádro) Je největší a zasahuje kaudálně až k jádrům zadních provazců míšních.
- boční předsíňové jádro – ncl. vestibularis lateralis (Deitersovo jádro)
- horní předsíňové jádro – ncl. vestibularis superior(cranialis) (Bechtěrevovo jádro)
- dolní předsíňové jádro – ncl. vestibularis inferior (caudalis) (Rollerovo jádro)
- Hlemýžďová jádra – ncll. cochleares. Leží zevně od jader vestibulárních v recessus lateralis fossae rhomboideae. Jádra přijímají první neuron sluchové dráhy, vycházející axony (efferentní vlákna) tvoří druhý neron sluchové dráhy a jako zevní klička- lemniscus lateralis končí v jádrech dolních hrbolků středomozkového krytu – colliculi inferiores tecti mesencephali:
- zadní hlemýžďové jádro – ncl. cochlearis posterior (dorsalis)
- přední hlemýžďové jádro – ncl cochlearis anterior (ventralis).
Střední mozek- mesencephalon:
Somatomotorická zóna:
- Jádro okohybného nervu – ncl. originis nervi oculomotorii (N III). Vydává efferentní vlákna od okohybných svalů (přímý prostřední, horní a dolní sval oční, dolní šikmý sval oční – m. rectus medialis, superior et inferior a m. obliquus inferior).
- Jádro kladkového nervu – n.originis nervi trochlearis (N VI). Vydává efferentní vlákna pro šikmý horní oční sval- m. obliquus superior.
Visceromotorická zóna:
- Párové parasympatické jádro Edinger-Westphalovo. Vydává vlákna pro ganglion ciliare a ty po přepojení pokračují (inervují) do svěrače zornice – m. sphincter pupilllae a svalu řasnatého – m. ciliaris.
Somatosenzitivní zóna:
- Středomozkové jádro trojklanného nervu – ncl. mesencephalicus N V. Je složeno z pseudounipolárních buněk. Afferetní vlákna přivádají propriocepci z čelistního kloubu, dásní, zubů a žvýkacích, mimických a okohybných svalů. Efferentní vlákna vedou do retikulární formace (FR), do motorických jader N V, a N VII a do mozečku.
- Horní předsíňové jádro – ncl. vestibularis superior (cranialis) (Bechtěrevovo jádro) Leží zevně od mencephalického trigeminového jádra, blíže k povrchu fossa rhomboidea.
- zevní popis
Síťovaná formace – formatio reticularis
vlastní struktura jádra
Je soubor šedých hmot, které se rozkládájí v celém kmeni mozkovém. Jde o systém jader a spojů „všeho se vším“, který vzájemným propojením vláken má siťovitý charakter. (rete = síť), proto se nazývá retikulární formace (RF). Axony RF propojují i značně vzdálené oblasti, jako je lůžko – thlamaus, mozeček a mícha, a tím aktivují mozek jako celek, mají dráždivý charakter, mluvíme o ascedentním budivém systému.
Jádra RF: jsou uspořádána ve třech podélných systémech:
- Švový systém – rapheální systém – nuclei raphe. Soubor nepárových jader ve střední čáře. Vydává vlákna převážně k buňkám mediálního systému RF a systému limbickému.
- Přístřední systém – mediální systém. Nejmohutnější soubor párových jader, uložených po obou stranách systému rapheálního. Jde o soubor velkých nervových buněk, ze kterých vychází většina spojů RF.
- Boční systém – laterální systém. Soubor drobnějších párových jader uložených po obou stranách mediálního systému, do kterého převážně vydává své axony.
Kromě těchto hlavních podélných systémů se v RF nacházejí skupiny buněk, které jsou charakterizované svým spojením nebo mediatorem.
Obecně se jádra RF uložená v prodloužené míše nazývají ncll. reticulares inferiores, uložená v pontu – ncll. reticulares medií a v mesencephalu – ncll. reticulares superiores.
Je to systém cerebellární a monoaminergní.
- Cerebellární systém vydává axony do mozečku
- Monoaminergní systém jsou buňky, které se dělí podle mediátoru na:
- Jádra katecholaminergní, jejich mediátorem je noradrenalin nebo dopamin, který je transportován do basálních ganglií a do mozkové kůry, a tím se významně podílí na činnosti aktivačního systému RF.
- Jádra serotoninergní, mediátorem buněk je serotonin.
- Jádra cholinergní, jádra jsou uložena v basální oblasti mozkových hemisfér a v mozkovém kmeni. Produkují acetylcholin. Buňky RF vydávají velké množství ascendentních a descendentních axonů a tím rozvádějí impulsy do blízkých i vzdálených struktur.
Afferentní dráhy (spoje RF):
- Míchosíťovaná dráha – tractus spinoreticularis. Přichází z jader zadních provazců míšních a má zkřížená i nezkřížená vlákna. Na nezkřížená vlákna navazují tractus reticulothalmicus – dráha siťovolůžková.. Tím je RF zapojena do systému senzitivity (převádí pomalou difuzní bolest a signály senzitivní).
- Jádrosiťová dráha – tractus nucleoreticularis. Vlákna přícházejí ze senzitivních jader hlavových nervů a jsou RF předány motorickým jádrům hlavových nervů – tractus reticulonuclearis – drána siťovojádrová. Tím je zajištěna funkce reflexu: senzitivní podnět- RF-motorická odpověď.
- Mozečkosiťová dráha – tractus cerebelloreticularis. Dráha vychází z části mozečku přes horní mozečkovou stopku – pedunculus cerebellaris superior do jader RF mesencephala (ncl. ruber) a jader pontu a přes dolní mozečkovou stopku – pedunculus cerebellaris inferior do jader RF míchy. Navazují na ně:
dráha siťovomíšní – tractus reticulospinalis, končí na míšních motoneuronech a dráha síťojádrová – tractus reticulonuclearis, končí na motorických jádrech hlavových nervů. Tyto dráhy převádějí vliv mozečku na míšní segmenty a tím je zajištěna kontrola pohybů mozečkem.
- Krytosíťovaná dráha – tractus tectoreticularis. Většina vláken je nezkřížených. Dráha vychází z hlubokých vrstev colliculi superiores a končí ve RF pontu a medullae oblongata. Navazuje na ni dráha síťovomíšní – tractus reticulospinalis.
- Podlůžkovosíťovaná dráha – tractus hypothalamoreticularis. Dráha sestupuje z jader hypothalamu a končí v rapheálních a mediálních jádrech RF. Navazuje na ni dráha síťojádrová – tractus reticulonuclearis, která končí na parasympatických jádrech hlavových nervů (NIII,VII,IX,X.) a dráha síťomíšní – tractus reticulospinalis, která končí na sympatickém jádru míchy (ncl.intermediolateralis).
- Bledosíťovaná dráha – tractus pallidoreticularis. Dráha vychází z basálních ganglií (bledé jádro – globus pallidus) a končí na jádrech RF pontu. Na RF má inhibiční vliv.
- Červenosíťová dráha – tractus rubroreticularis a
- Černosíťová dráha – tractus nigroreticulairs. Dráhy vycházejí z příslušných jader a končí na jádrech RF pontu. Efferentní vlákna vydávají dráhy do okruhu basálních ganglií, telencephala a také jako tractus reticulospinalis končí na motorických jádrech míchy. Na RF mají opět inhibiční vliv a účastní se na řízení a regulaci motoriky.
- Korosíťová dráha – tractus corticoreticularis. Dráha vychází z motorických i senzitivních korových oblastí mozku a končí na všech systémech jader RF. Na ni navazují 3 významné dráhy: tractus reticulospinalis, tractus reticulonuclearis a tractus reticulocerebellaris. Tím je zajištěna korová kontrola motoriky.
- Další kontrola motoriky je zajištěna napojením RF na thalamus, subthalamus a mandlové jádrao (ncl. amygdaloideum).
Efferentní dráhy (spoje RF):
Jsou převážně dlouhé spoje vycházející z mediálního systému RF a končí (rozvádějí signály) ve vzdálených strukturách a oblastech CNS. Některé jejich funkce byly opakovaně objasněny u afferetních spojů RF.
- Dráha síťomíšní – tractus reticulospinalis. Obsahuje vlákna vycházející převážně z jader v pontu a medula oblongata. Dráha sestupuje předními míšními provazce a končí na motorických jádrech míchy. Navazuje na dráhy:
tr.corticoreticularis, tr.cerebelloreticularis, tr. hypothalamoreticularis a mnoho dalších afferentních spojů RF.
- Dráha síťojádrová – tractus reticulonuclearis. Vychází z jaderRF a knočí na senzitivních a motorických jádrech hlavových nervů. Přepojuje signál z tractus nucleoreticularis, tractus hypothalamoreticularis a tractus corticoreticulairs. Motorická jádra hlavových nervů aktivují, senzitivní jádra inhibují.
- Síťosíťovaná dráha – tractus reticuloreticularis. Spojuje navzájem buňky retikulární formace krátkými i dlouhými axony. Plní aktivační funkci RF.
- Dráha síťomozečková – tractus reticulocerebellaris. Z buněk RF jsou do mozečku přiváděny signály z míchy, vestibulárního aparátu a mozkové kůry. Tím je zajištěna mozečková kontrola pohybů.
- Dráha siťohrbolová (síťolůžková) – tractus reticulothalamicus. Vlákna vystupují hlavně z jader mediálního systému RF a cholinergních jader RF a vedou vzruchy do intralaminárních jader a předních jader thalamu. Axony – efferetní vlákna jsou hlavní složkou aktivačního systému RF, jsou i součástí přenosu signálů bolesti.
- Dráha síťopodhrbolová (siťopodlůžková) – tractus reticulohypothalamicus. Vystupuje hlavně z rapheálních jader RF a knočí na jádrech hypothalamu, většina vláken je serotoninergních. Ovlivňuje činnost nejvyšších vegetativních center, činnost hypothalamo-hypofyzárního systému a ovlivňuje i termoregulační funkce.
Funkce retikulární formace
RF má vlké množství aferetních, eferetních a intraretikulárních spojů a tím může plnit velké množství funkcí. Jsou to funkce aktivační, inhibiční, spojovací a koordinační.
Ascendentně jsou dráhy RF zapojeny hlavně do systémů senzitivity (vedení bolesti) a aktivačních systémů CNS (udržování vědomí) – RAS – retikulární ascendentní systém. Zrakové, sluchové a hmatové stimuly nás udržují v bdělosti a duševně ve střehu. RAS také funguje během spánku a při probuzení. Naopak alkohol, sedativa, celkové anestezie, které RAS potlačují mohou vést ke ztrátě bdělosti a vědomí. Při vážném poranění dojde ke komatu (údery při boxu apod..)
Descendentně jsou dráhy RF zapojeny hlavně do motoriky a vegetativních spojů (připojení drah z hypothlamu na sympatikus a parasympatikus). Některé eferetní spoje RF fungují aktivačně – facilitačně, jiné inhibičně. Na střídavé souhře facilitace s inhibici je založena i činnost některých retikulárních center (centra RF) (dýchací, vasomotorické, regulace srdeční činnosti).
RF plní i funkci mnohočetných reflexů a životně důležitých center:
Obživné reflexy: reflex polykací, sací, slinivý, visceromotorický (sekrece šťáv a pohyby GIT cestou vegetativních nervů)
Obranné reflexy: reflex slzivý, mrkací, kašlací, dávivý, zornicový.
Dýchací centrum: má inhibiční a expirační oddíl. Cestou tractus reticulospinalis jdou vlákna k motoneuronům hlavních i pomocných dýchacích svalů a zajišťují jejich regulaci.
Pneumatické centrum. Je nadřazeno centru dýchacímu. Působí na něho řada vlivů: informace o hladině O2(oxygenaci), hladině CO2 (kapnii) v krvi, podněty z napětí svalů a šlach, podněty z vyšších úseků CNS (automatické ovlivnění dýchání – bolest, strach, emoce…., reflexní ovlivnění dýchání – kýchání, kašel, zívání, řeč, zpěv, TK, tělesná teplota a některé hormony).
Centrum pro regulaci srdeční činnosti. Je nadřazeno vegetativnímu systému (sympatiku a parasympatiku) a má kardioakcelerační a kardioinhibiční funkci.
Vasomotorické centrum. Je taktéž nadřazeno vegetativnímu systému, reguluje TK a tím dále ovládá kontrakce a dilatace arteriol a prekapilár.
Centrum zvracení. Je napojeno na jádra N X. a osamocené jádro – ncl. solitarius, které do centra přivádějí podněty z chemoreceptorů GIT a dýchacích cest.
Regulace tělesné teploty: zajišťuje ji podhrbolosíťovaná dráha – tractus hypothalamoreticularis, která končí na rapheálních jádrech RF.
Funkce RF při vedení bolesti: zajišťuje tractus spinoreticulothalamocorticalis. V mozkové kůře dochází k vnímání pomalé, difuzní bolesti.
| Obrázky ke kapitole: Mozkový kmen – zadní mozek | |||
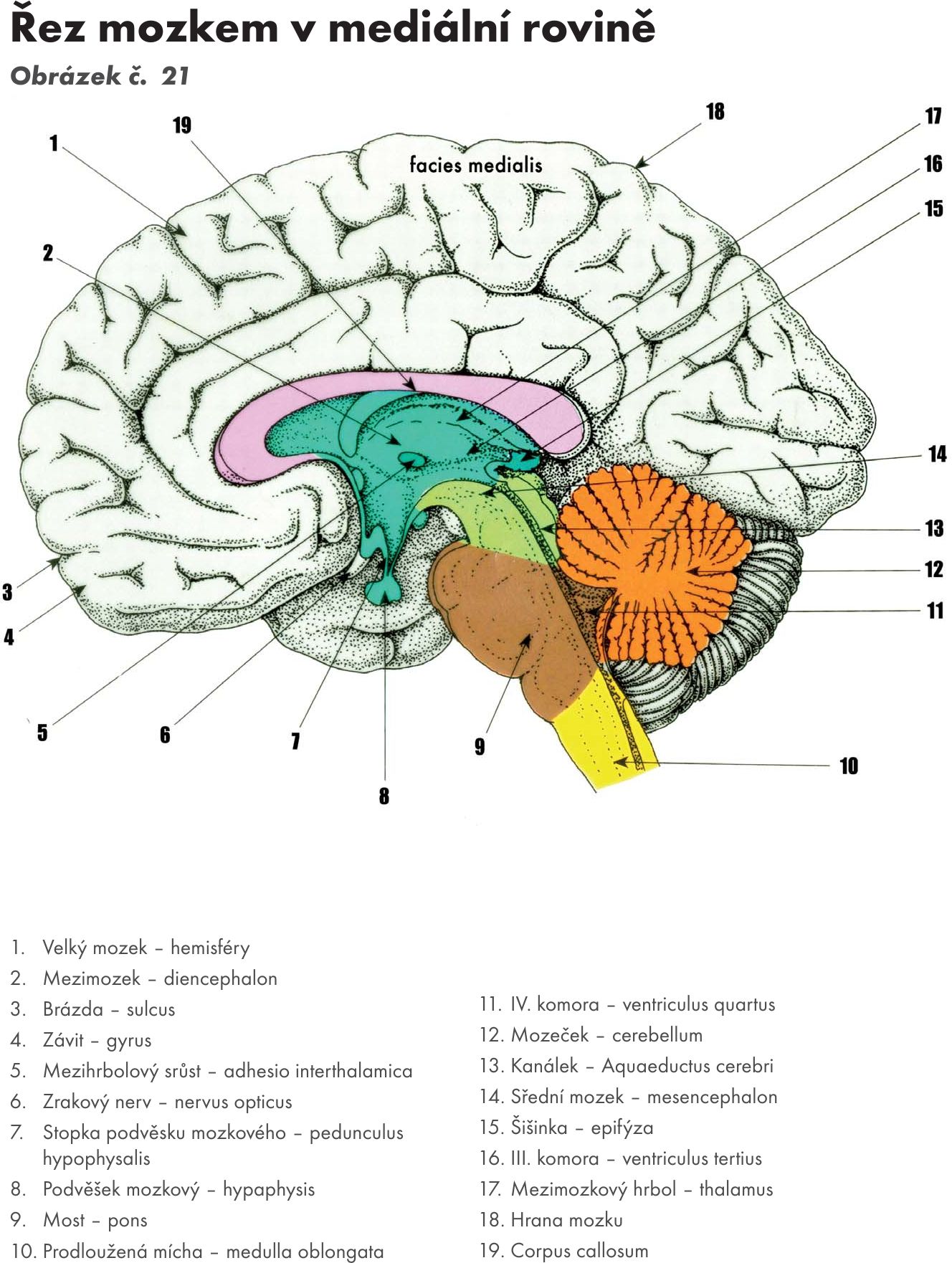
Obr. 21 Řez mozkem v mediální rovině
|
Obr. 22 Struktura prodloužené míchy
|
Obr. 23 Struktura prodloužené míchy
|

Obr. 24 Mozkový kmen – Varolův most
|
Obr. 25 Mozkový kmen
|
Obr. 26 Struktura Varolova mostu
|
Obr. 27 Struktura Varolova mostu
|
Obr. 28 Struktura Varolova mostu
|
Obr. 29 Spodina mozková a výstupy hlavových nervů
|
Obr. 30 Structura středního mozku – mesencephala
|

Obr. 31 Struktura mesencephala
|
Obr. 32 Kosočtvercová jáma (spodina IV. komory)
|
Mozeček – cerebellum
-
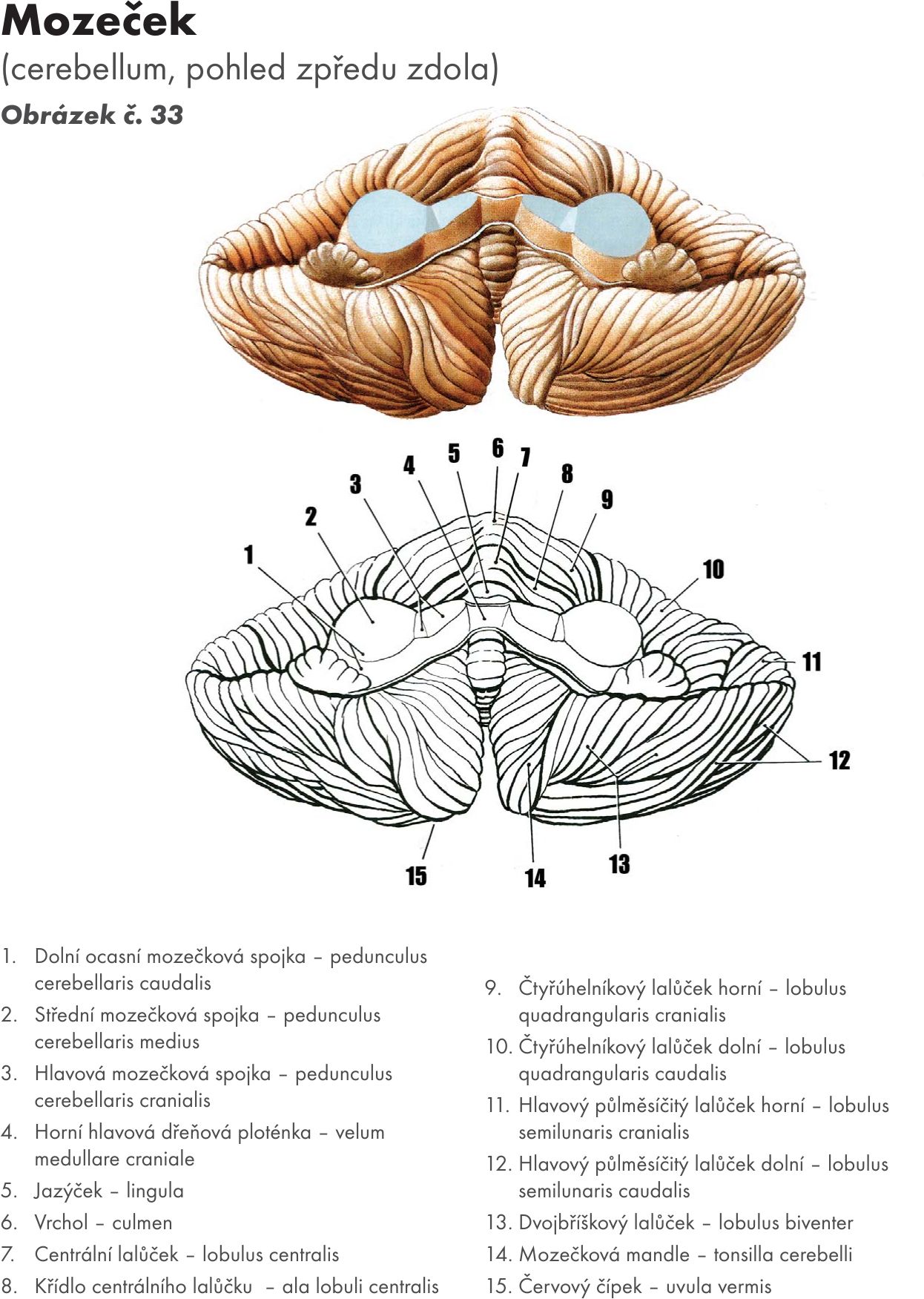
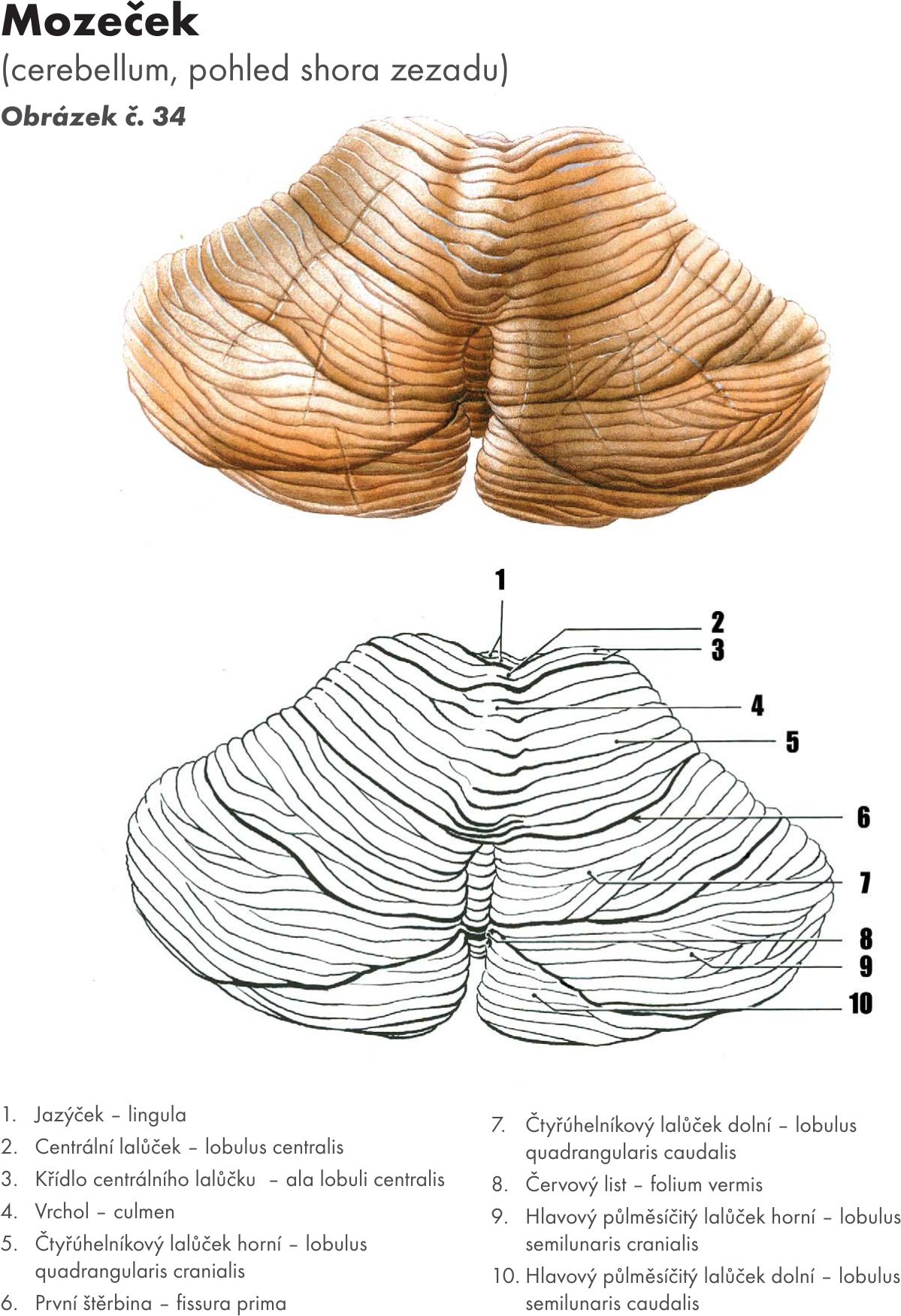
Zevní popis Obrázek č. 33 Obrázek č. 34
Mozeček leží v zadní jámě lebeční a od týlního laloku je oddělen derivátem tvrdé pleny mozkové- napjatá blána – tentorium cerebelli. Má květákovitý tvar a tvoří 11 % hmoty mozku. Je umíštěn nad IV.komorou, která vybíhá proti mozečku cípem – fastigium. Váha mozečku je asi 130–150 g.
Mozeček se skládá ze střední oblasti – mozečkový červ – vermis cerebelli, kolem je paravermální zóna a laterálně jsou uloženy dvě polokoule mozečkové – hemisheriae cerebelli.
S mozkovým kmenem je mozeček spojen třemi páry mozečkových stopek, které tvoří svazky bílé hmoty. S prodlouženou míchu to jsou dolní mozečkové stopky – pedunculi cerebelli inferiores – corpora restiformia, s Varolovým mostem střední mozečkové stopky – pedunculi cerebelli medii – brachia pontis a se středním mozkem horní mozečkové stopky – pedunculi cerebelli superiores – brachia conjunctiva.
Povrch mozečku je rozdělen dvěma hlubokými rýhami na tři laloky (lobi), které jsou dalšími štěrbinami (fissurae) členěny na lalůčky (lobuli) a ty jsou mělkými rýchami (sulci) členeny na mozečkové lístky (folia). Tímto čelenením tvoří povrch kůry mozečku až 40 % povrchu hemisfér velkého mozku.
Laloky se dělí podle vývojového hlediska :
- Vločkouzlíkový lalok – lobus flocculonodularis. Je vývojově nejstarší část, tzv. pramozeček (předsíňový mozeček) – archicerebellum. Má dvě části: uzlík – nodulus v oblasti mozečkového červu – vermis a vločku – flocculus v oblasti hemisfér. Afferetní vlákna přijímá převážně z vestibulárních jader, proto se také nazývá vestibulární mozeček – vestibulocerebellum. Jeho funkcí je správné držení těla.
- Hlavový (přední) lalok – lobus cranialis. Je vývojově mladší tzv. starý mozeček (míšní mozeček) – paleocerebellum. Afferetní vlákna přijímá převážně z míšních jader, proto se také nazývá míšní mozeček – spinocerebellum.
- Ocasní (zadní) mozeček – lobus caudalis. Je vývojově nejmladší, tzv. nový mozeček (mozkový mozeček) – neocerebellum. Afferetní vlákna přijímá z kůry přes pons (ncl. pontis), proto se také nazývá nový mostní mozeček – pontocerebellum.
Na povrchu mozečku je tenká vrstva šedé hmoty – mozečková kůra – cortex cerebelli, pod ní je bílá hmoty, která vybíhá do jednotlivých mozečkových lístků – folia cerebelli. Toto stromečkovité větvení se označuje mozečkový strom života – arbor vitae cerebelli.
Kůra mozečku se skládá ze tří vrstev: Obrázek č. 35
Molekulární vrstva (pleteňovitá)(cinereum) – stratum moleculare (plexiformis). Je povrchová vrstva kůry a obsahuje dva typy buněk
- hvězdicovité buňky, jsou uloženy v povrchových oblastech vrstvy, a
- košíčkovité buňky, jsou uloženy hlouběji.
Dendrity a axony obou buněk probíhají paralelně s povrchem folií a jejich aoxony končí na dendritech Purkyňových buněk druhé vrstvy mozečkové kůry. Na Purkyňovy buňky mají inhibiční vliv. Do molekulární vrsty zasahují dendrity Purkyňových buněk a axony granulačních buněk z třetí vrstvy mozečkové kůry. Tyto se dělí ve tvaru písmene T na dvě kolaterály, které probíhají paralelně s povrchem kůry. Mají synaptické kontakty s dendrity Purkyňových, hvězdicovitých i košíčkových buněk a jejich axony působí aktivačně. Do stratum moleculare přicházejí i šplhavá vlákna z olivárních jader. Procházejí (šplhají) dvěmi hlubšími vrstvami a vždy jedno vlákno se větví na dendritech jedné Purkyňovy buňky. Šplhavá vlákna působí aktivačně.
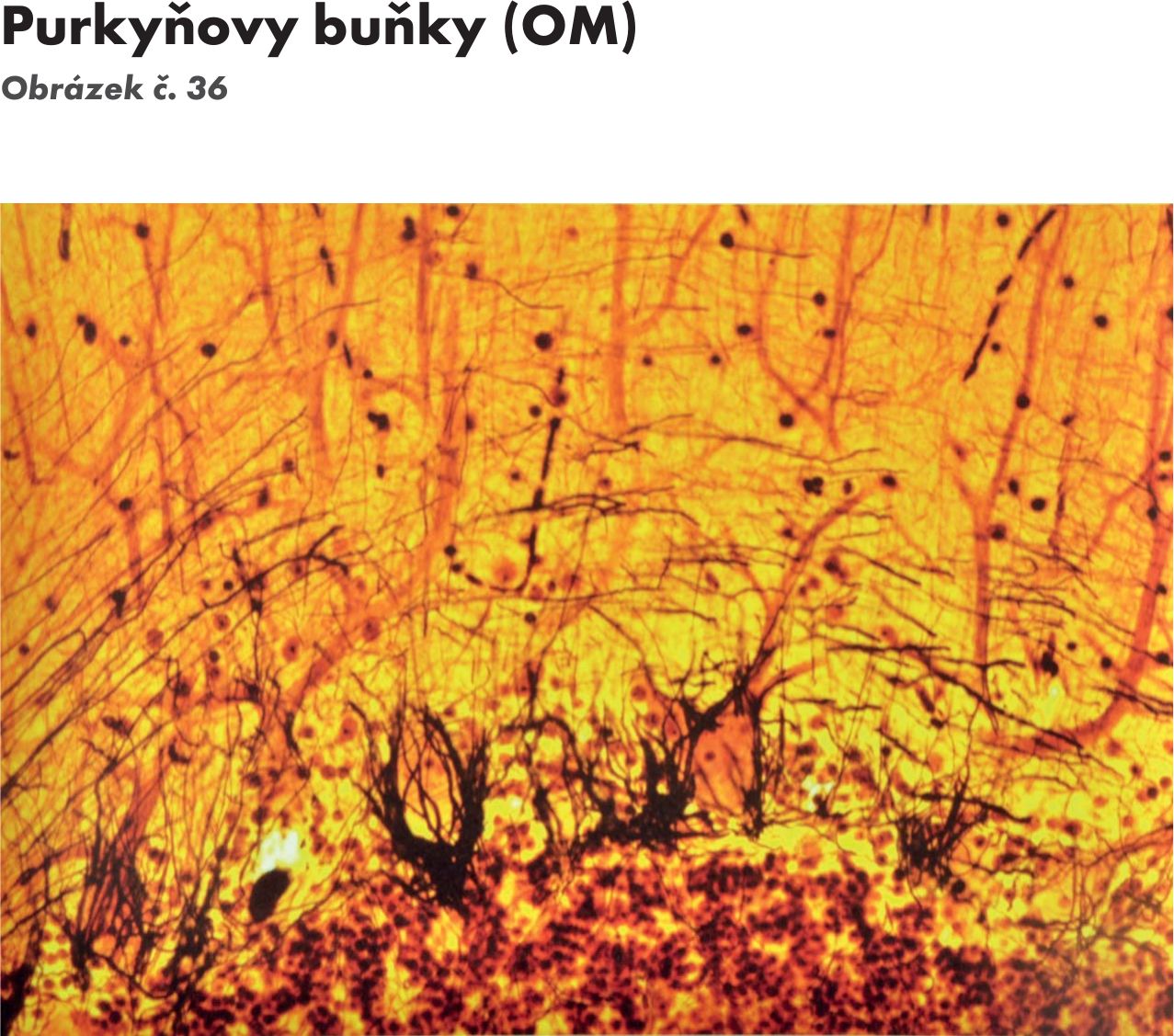
- Gangliová vrstva – stratum gangliosum. Je střední vrstva mozečkové kůry. Obsahuje velké množství velkých neuronů (50–60μm) hruškovitého tvaru – tzv. Purkyňovy buňky Obrázek č. 36. Jejich dendrity vstupují do molekulární vrstvy, kde vytvářejí bohatou dendritickou arborizaci orientovanou kolmo na povrch kůry (C.Golgi). Jejich axony jsou myelinizované, tvoří bílou hmotu mozečku, a končí u jader mozečku. Jsou inhibiční a jejich mediátorem je GABA (kyselina γ- aminomáselná). Vrstvou procházejí šplhavá vlákna.
- Zrnitá vrstva – stratum granulare (ferrugineum – na čerstvém řezu má slabě narezlou barvu). Je hluboká vrstva mozečkové kůry. Obsahuje granulační (zrnité) buňky a buňky Golgiho.
- Granulární buňky. Jsou nejmenší nervové buňky CNS. Jejich krátké dendrity vrstupují do mozečkových glomerulů. Axony granulačních buněk stoupají do molekulární vrstvy, kde končí jako paralelní vlákna (rovnoběžná s povrchem kůry), mají aktivační vliv a jejich mediátorem je glutamát.
- Golgiho buňky. Jejich dendrity zasahují do všech vrstev mozečku, jejich axony, tak jako dendrity granulačních buněk, vstupují do mozečkových glomerulů.
Báňky hvězdicovité, košíčkovité, granulační(zrnité) i Golgiho jsou interneurony, jejich axony neopouštějí mozečkovou kůru. Pouze axony Purkyňových buněk po přepojení na mozečkových jádrech vycházejí z mozečku jako projekční drýhy. Kromě granulačních buněk jsou všechny ostatní buňky mozečkové kůry inhibiční, což je důležité pro průběžnou kontrolu všech probíhajících pohybů.
Mozečkový glomerulus. Je složitý synaptický komplex uložený mezi granulačními buňkami. Přichází do něho presynaptické zakončení mechového vlákna, které je zde v kontaktu s dendrity většího počtu granulačních buněk (10–20), na které mají aktivační vliv. V glomerulu s dendrity granulačních buněk mají kontakt i axony Golgiho buněk, které mají inhibiční vliv.
-
Vlákna mozečkové kůry:
- Mechová vlákna. Jsou axony z míšních jader zadních provazců (ncl. gracilis a ncl. cuneatus) a z jader hlavových nervů. Končí v mozečkovém glomerulu v granulační vrstvě, zde jsou v kontaktu s dendrity většího počtu granulačních buněk. Vlákna působí aktivačně.
- Šplhavá vlákna. Jsou axony z olivárnách jader (ncl olivaris inferior), procházejí až do molekulární vrstvy, kde mají synaptické kontakty s dendrity Purkyňových buněk – vždy jedno vlákno s dendritem jedné Purkyňovy buňky. Vlákna působí aktivačně.
- Multilaminární vlákna. Přicházejí z monoaminergního systému RF a větví se ve všech vrstvách mozečkové kůry.
Všechny tří typy vláken jsou převážně myelinizované, do mozečku se dostávají cestou mozečkových stopek (pedunculi cerebellares). Spolu s axony Purkyňových buněk, které končí na mozečkových jádrech a z nich vystupující dráhy (vlákna), které končí na jádrech mozkového kmene a thalamu, tvoří bílou hmotu mozečku.
Mozečková jádra : Obrázek č. 37
Jsou čtyři jádra zanořená do bílé hmoty mozečku a nazývají se podle tvaru nebo uložení. Vstupují do nich afferetní vlákna – axony Purkyňových buněk a kolaterály mechových a šplhavých vláken.
- Zubaté jádro – nucleus dentatus. Je největší mozečkové jádro, v hemisféře leží nejlaterálněji a jeho branka- hilus se otvírá mediálně a rostrálně (kraniálně). Odpovídá neocerebellu.
- Zátkovité jádro – nucleus emboliformis. Je menší jádro, které mediálně přiléhá k hilu zubatého jádra – ncl. dentatus.
- Kulovité jádro – nucleus globosus. Je tvořeno více kulatými buněčnými skupinami a leží mediálně od zátkovitého jádra.
- Cípovité jádro – nucleus fastigii
Ncl. emboliformis a globosi odpovídají paleocerebellu.
Na buňky mozečkových jader působí axony Purkyňových buněk inhibičně, kolaterály mechových a šplhavých vláken aktivačně.
-
Spoje mozečku:
Afferetní spoje. Obrázek č. 38 Obrázek č. 39
Archicerebellum: dráhy přicházejí z vestibulárních jader cestou pedunculi cerebellares inferiores do vločkouzlíkového laloku – lobus flocculonodularis (ncl fastigii). Do mozečku přicházejí signály o poloze a pohybech hlavy. (tr. vestibulocerebellaris).
Paleocerebellum : dráhy přicházejí z jader míšních, bulbárních, RF, olivárnách, ze senzitivních jader N V., z osamocené jádro – ncl. solitarius, a jader hypothalamu. Končí na zátkovitém a kulovitém jádru (ncl. emboliformis et globosus).
Neocerebellum : dráhy přicházejí ze široké oblasti mozkové kůry (z motorických, senzitivních a ze zrakové kůry) a po přepojení na ncl pontis nebo bez přepojení končí na zubatém jádru – ncl. dentatus. (tr. corticopontocerebellaris). Přivádejí informace o aktivitě neuronů rozsáhlých korových oblastí a informace související s přípravou a provedením pohybů. Dráhy přicházejí cestou středních mozečkových stopek – pedunculi cerebellares medii.
Efferetní spoje. Obrázek č. 40
- Spoje z kůry mozečku do mozečkových jader – kortikonukleární spoje. Zajišťují axony Purkyňových buněk ve třech podélných zónách:
- mediální zóna do ncl fastigii
- paramediánní zóna do ncl. globosus a ncl. emboliformis
- laterální zóna do ncl dentatus.
Na neurony mozečkových jader mají Purkyňovy buňky inhibiční vliv.
- Spoje z nucleus fastigii (z vestibulárního mozečku). Vlákna končí na jádrech vestibulárních a jádrech RF pontu a medulla oblongata. Na ně navazují vlákna (dráhy), která jdou na motorická jádra míšní a mají excitační vliv na motoneurony extenzorů (horních končetin, dolních končetin, šíjové a zádové svaly).
- Spoje z nucleus emboliformis a nucleus globosi (ze spinálního mozečku). Vlákna končí na jádrech thalamu, RF, ncl. ruber a jádrech olivárnách. Na ně se napojují dráhy do jader vyšších i nižších etáží CNS.
- Spoje z nucleus dentatus. Vlákna končí na jádrech: ncl ruber, ncl interstitialis Cajali, ncl Dakschewitschi, ncl. N III a jádrech thalamu. Navazujícími drahami ovlivňuje ncl. dentatus aktivitu korových neuronů a motoneurony inervující distální svaly končetin.
- Spoje z kůry mozečku do mozečkových jader – kortikonukleární spoje. Zajišťují axony Purkyňových buněk ve třech podélných zónách:
-
Funkce mozečku Obrázek č. 41
Mozečková kůra koordinuje pohyby těla, je důležitým regulačním centrem motoriky. K tomu potřebuje příjem tří typů informací:
- Informace o rovnováze. Získává z receptorů vnitřního ucha přes vestibulární jádra.
- Informace o aktuálních pohybech končetin, krku a trupu. Získává z proprioreceptorů uložených ve svalech, šlachách a kloubech. Do mozečku se dostávají přes afferetní dráhy spinálního mozečku (paleocerebellum).
- Informace z mozkové kůry. Tyto se do mozečku dostávají pyramidovou dráhou a jádry ve Varolově mostě (ncl. pontini).
Na základě těchto informací
- Zabezpečuje udržování rovnováhy. Zodpovídá především vestibulární mozeček. Při poškození se objevují poruchy stoje a chůze.
- Reguluje svalový tonus a reflexní dráždivost. Zodpodívá především spinální mozeček. Působí tlumivě na antigravitační svalstvo. Při poškození dochází ke zvýšení tonu extenzorů.
- Zajišťuje svalovou koordinaci pohybu. Uplatňuje se především pontinní mozeček. Přijímá informace z mozkové kůry o přípravě pohybu. O vlastní realizaci pohybu získává signály z proprio- a extero-receptorů. Ty porovnává s probíhajícími pohyby těla a zpětně vysílá instrukce do mozkové kůry, tím mozeček průběžně srovnává korovou a periferní informaci. Použitím této zpětné vazby z mozečku může mozková kůra znovu opravovat motorické povely a posílat je k míše. Výsledken jsou sladěné a dobře koordinované pohyby.
Při poškození dochází k neschopnosti provádět střídavě opačné pohyby, objevují se i třes při pohybu, poruchy řeči. Trajektorie pohybu není přímá, ale připomíná Brawnův pohyb.
Při krvácení nebo nádorech mozečku se často mohou objevit příznaky z poškození tří funkčních částí mozečku.
Při mozečkových postiženích bývá narušeno harmonické provádění volních pohybů,včetně jejich rychlosti.Je narušena i automatická koordinace.
Příznaky:
- Dyssynergie – chybí koordinace různých pohybů zúčastněných svalů a svalových skupin (záklon trupu bez současného ohnutí kolen, ohrožující rovnováhu; při chůzi po čtyřech nejsou končetiny používány střídavě přesně do kříže).
- Dysmetrie – chybí míra pro přiměřený rozsah a rychlost volního pohybu (přehnané rozevírání prstů ve snaze o uchopení drobného předmětu; snaží-li člověk při zavřených očích vystoupit na lavici,zvedá nohy příliš vysoko).
- Ataxie – všechny svaly při provádění volního pohybu nepracují současně a harmonicky k optimálnímu dosažení cíle (testy: prst-nos, pata-koleno-holeň – s předozadní výchylkou končetiny z ideálního směru pohybu). Ataxie trupu v sedě
- Intenční třes – při pohybu dochází ke vzniku třesu pohybující se části těla.
- Patologický rebound fenomén – antagonisté selžou při včasném zbrždění přestřelujícího pohybu, když např. flexory lokte, jsou- li kontrahovány, i po náhlém odporu ruky vyšetřujícícho, pokračují v kontrakci i po náhlém odtažení této ruky, anebo pokus s oboustranně dopředu nataženými pažemi tlačícími proti tlaku vyšetřujícího.
- Dysdiadochokinéza – rychle alterující pohyby agonistů a antagonistů nejsou již dostatečně rychlé a plynulé: rychlá pronace a supinace předloktí, rychlé poklepávání stehna střídavě dlaní a hřbetem vlastní ruky.
- Hypotonie – v průběhu pasivních pohybů (pasivní třepání končetin, silné kmitání rukou stojící osoby, kterou je prostřednictví ramen otáčeno kolem jeho tělesné osy dopředu a dozadu).
- Pokles v pozičním pokusu – pacientovo svalové napětí je nedostatečné k udržení přiměřených postojů, a tak jsou-li zavřené oči, natažená paže na straně postižení klesá.
- Deviace v Barányiho ukazovacím pokusu – zvedne-li pacient ruku a zacílí-li prst na bod před sebou, paže na straně postižení při zavřených očích poklesne. Na postižené straně má tendenci se uchylovat ke straně postižení.
- Nejistota postoje
- Nejistá chůze o široké bázi, je popisována osobou jako závrať, avšak nepředstavuje pravé rotatorní vertigo.
- Nystagmus – při mozečkových lezích bývá přítomen hrubý nystagmus ke straně postižení, který se zvýrazňuje pohledem k postižené straně a mizí při zavřených očích.
- Porucha řeči – vyrážená, explozivní řeč.
Zastoupení jednotlivých částí těla v mozečku je topické. Konkrétní mozečkové regiony mají rovněž konkrétní funkce. Lze tak orientačně přisuzovat k jednotlivým příznakům lokalizační význam:
- V případě bazálních a ve střední linii uložených procesů: poruchy držení trupu a rovnováhy, zvláště při sezení
- Při uložení procesů v rostrální části střední linie: poruchy koordinace postihující stoj i chůzi.
- Při procesech v laterálních oddílech (hemisférách): porucha koordinace pohybů (zručnosti) homolaterálních končetin.
| Obrázky ke kapitole: Mozeček | |||
Obr. 33 Mozeček
|
Obr. 34 Mozeček
|

Obr. 35 Kůra mozečku
|
Obr. 36 Purkyňovy buňky (OM)
|

Obr. 37 Horizontální řez mozečkem, jádra mozečku
|

Obr. 38 Schéma přívodních drah mozečku z míchy a jader mozkového kmene
|
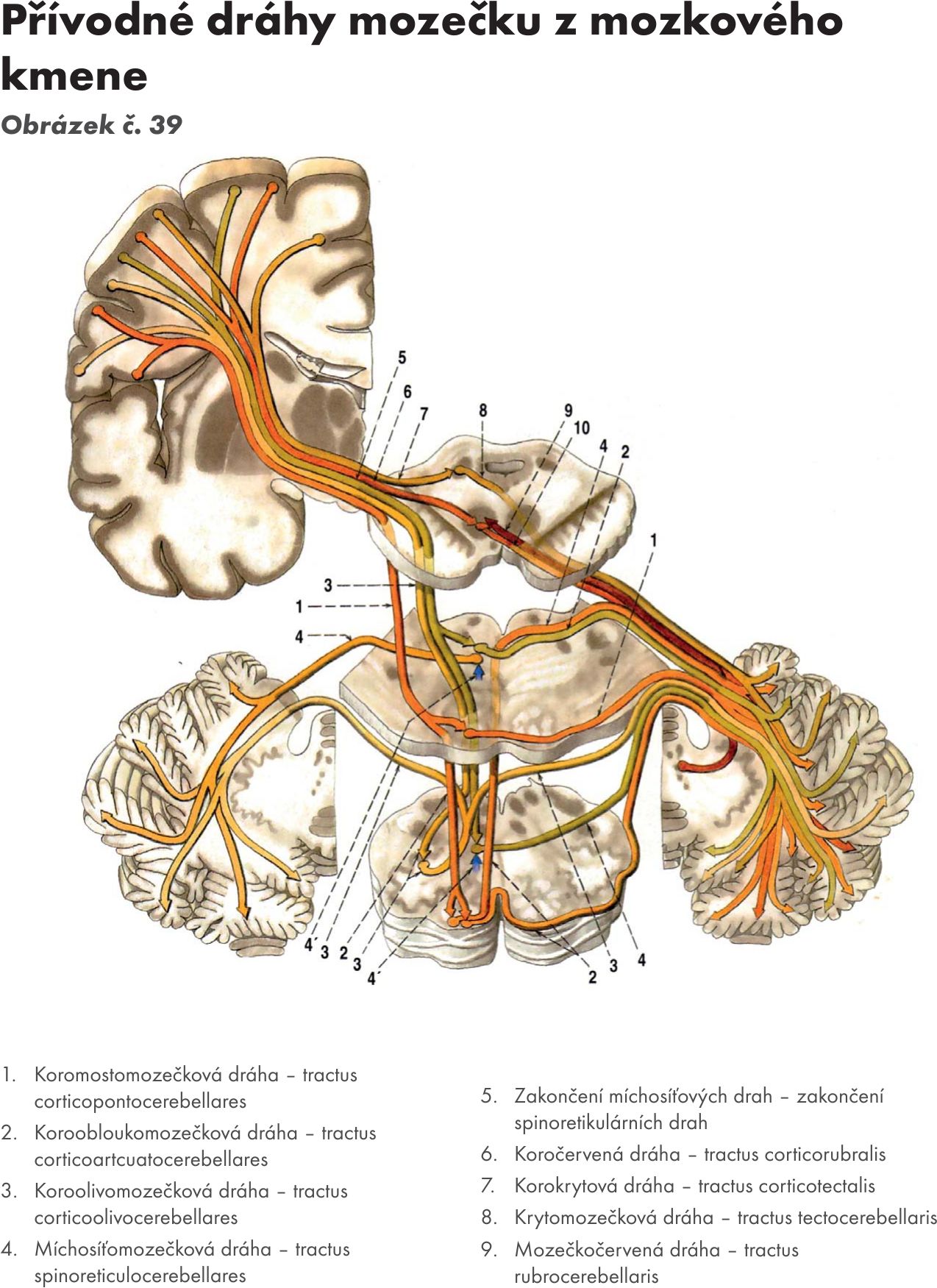
Obr.39 Přívodné dráhy mozečku z mozkového kmene
|
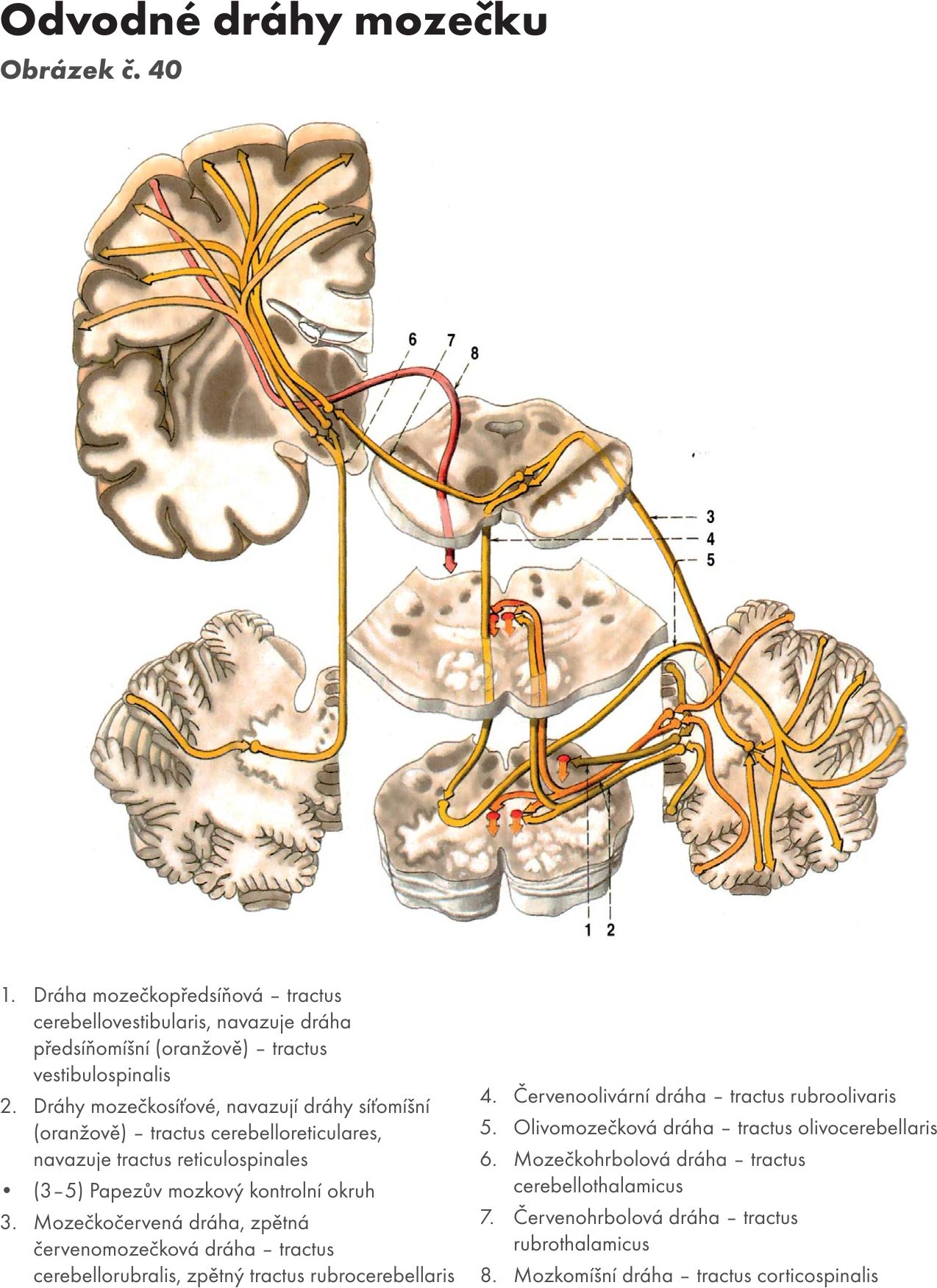
Obr. 40 Odvodné dráhy mozečku
|
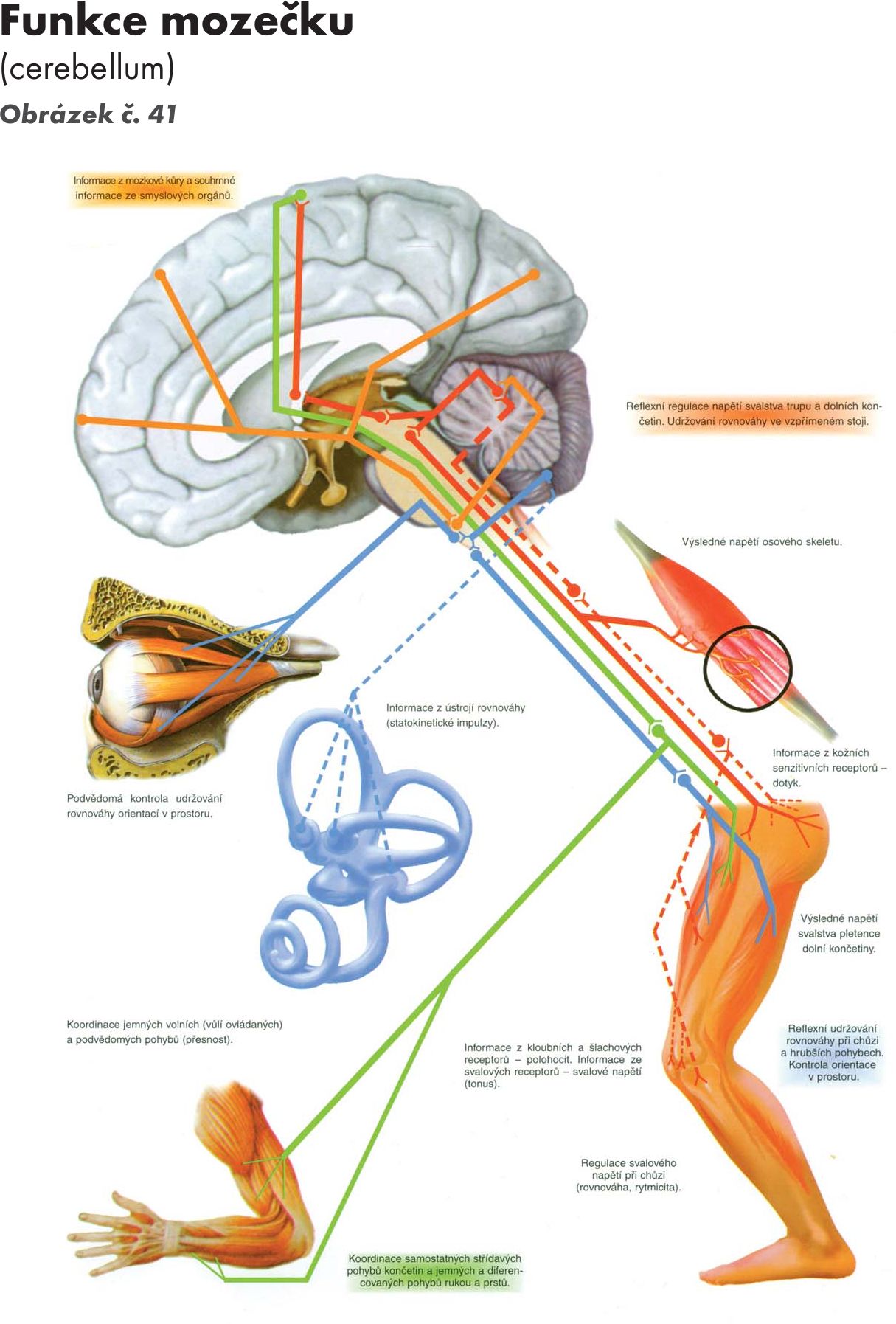
Obr. 41 Funkce mozečku
|
|||
Mezimozek – diencephalon
Skladá se z :
- hrbolu mezimozkového – thalamu,
- podhrbolí – hypothalamu,
- dolního hrbolu, předního hrbolu mezimozkového – subthalamu,
- nadhrbolí – epithalamu a
- záhrbolí – metathalamu.
-
Thalamus – hrbol mezimozkový
Thalamus je párový orgán, vejčitého tvaru, který se vpředu zužuje v přední hrbolový hrbolek – tuberculum anterius thalami a vzadu se rozšiřuje v polštář – pulvinar. Mediální plochy thalamu ohraničují laterálně III. komoru mozkovou, od podhrbolové brázdy – sulcus hypothalamicus (odděluje thalamus od podhrbolí – hypothalamus), k hrbolovému dřeňovému proužku – stria medullaris thalami mezi kterými se upíná strop 3. komory mozkové, který je tvořen ependymem. Strop srůstá s měkkou plenou – pia mater a cévními pleteněmi a vzniká cévnatá tkáň 3. komory – tela chorioidea ventriculi tertii. 3. komorou prochází mezihrbolový srůst – adhesio interthalamica, který obsahuje glii, šedou i bílou hmotu. Na dorsální ploše hrbolu – thalamu je laterálně patrný hraniční proužek – stria terminalis, který hrbol – thalamus odděluje od ocasatého jádra – nucleus caudatus (patří k basálním gangliím, str…). Bazálně od ocasatého jádra – ncl. caudatus srůstá s laterální plocha hrbolu – thalamu s bílou hmotou konečného mozku – telencephala. Dole srůstá s podhrbolím – hypothalamem a dolním hrbolem, předním hrbolem mezimozkovým – subthalamus.
Mikroskopická stavba
Hrbol – thalamus tvoří šedá hmota, kterou lze rozdělít průběhem plotének bílé hmoty na celou řadu jader. V thalamu se setkáváme se třemi typy neuronů (projekční, inhibiční a specifický typ neuronů v nucleus reticularis thalami), které rozlišujeme na základě rozdílů v morfologii, mediátorech, které jsou v nich exprimovány a jejich projekcí do ostatních oblastí nervového systému.
- Projekční neurony – projikují do kůry či striata, jedná se o neurony excitační, jejich mediátorem je glutamát.
- Inhibiční interneurony – inhibují projekční neurony, mediátorem je GABA.
- Neurony nc. reticularis thalami – inhibují ostatní thalamická jádra, jejich mediátorem je také GABA.
Jádra jsou pojmenována dle lokalizace.
I. dělení – dle topografického uložení
Thalamická jádra jsou oddělena ploténkami (laminae medullares thalami externa et interna), čímž vytváří morfologicky sedm skupin jader:
- Přední skupina jader je tvořena nc. anterodorsalis, nc. anteroventralis, nc. anteromedialis. Spoje k jádrům přicházejí z corpora mammillaria a sama vysílají vlákna do cingulární kůry velkého mozku.
- Jádra střední linie (nuclei mediani) jsou uložená pod okrajem III. mozkové komory. Patří sem ncc. paraventriculares anteriores et posteriores, nc. rhomboidalis, nc. reuniens.
- Mediální jádra – řadíme sem velké nc. medialis dorsalis.
- Nuclei intralaminares – jsou skupiny neuronů vnořených do lamina interna. Patří sem nc. paracentralis, nc. parafascicularis, nc. centralis lateralis, nc. centralis medialis, nc. centromedianus. Z nich vycházejí vlákna do čelní mozkové kůry.
- Ventrolaterální jádra – jsou největší skupinou thalamických jader. Řadíme sem nc. ventralis anterior (aferentace přichází z nc. globosus, eferentace míří ke striatu), nc. ventralis medialis (aferentace z mozečku, eferentace do motorických oblastí mozkové kůry), nc. lateralis dorsalis a nc. lateralis posterior (obě mají eferentace do kůry temenního laloku a limbické kůry), nc. ventralis lateralis (eferentace do motorické kůry I a II se somatotopickým uspořádáním), ncc. ventrales posteriores neboli tzv. ventrobazální komplex thalamu, kde jsou dvě jádra: nc. ventralis posterolateralis (aferentace z mediálního lemnisku a ze spinothalamických drah) a nc. ventralis posteromedialis (aferentace ze senzorických složek hlavových nervů, eferentace do somatosenzorické kůry postcentrálního závitu).
- Zadní skupina jader (nuclei posteriores) – zahrnuje asociační jádro pulvinar (eferantace z něj vedou do temenní a spánkové kůry) a dvě důležitá přepojovací jádra:
- zrakové dráhy – nc. corporis geniculati lateralis (pars dorsalis)
- sluchové dráhy – nc. corporis geniculati medialis (pars dorsalis).
- Retikulární jádra – tvoří tenkou vrstvu při vnější ploše. Jedná se o pokračování nespecifického ascendentního aktivačního systému retikulární formace mozkového kmene.
II. dělení – dle funkce
Jádra se mohou rozlišovat podle toho, zda je jejich funkce specifická – senzorická nebo nesenzorická, asociační nebo nespecifická. Toto dělení je však nutno brát s rezervou, proto je výhodnější vyhnout se přesnému kategorizování.
V thalamu lze nalézt tyto funkční skupiny jader:
- Jádra převádějící vzruchy z periferie do senzorických oblastí mozkové kůry.
- Jádra převádějící vzruchy z mozečku do motorické kůry.
- Jádra převádějící vzruchy z mezimozku do limbické kůry.
- Jádra zajišťující propojení jader mezimozku s asociačními oblastmi mozkové kůry.
- Jádra, která vysílají široké difůzní projekce, především do korových oblastí.
Podle volby parametrů můžeme mezi specifická jádra zařadit skupiny 1–2 nebo 1–3, mezi asociační skupinu 4 a do nespecifických jader skupinu 5.
- Specifická senzorická – somatosenzorická jádra – sem patří lemnisková jádra nc. ventralis posterolateralis, nc ventralis posteromedialis (pro oblast hlavy), nc. corporis geniculati medialis (pars dorsalis) je podkorovým převodním jádrem sluchové dráhy, obdobně funguje nc. corporis geniculati lateralis (pars dorsalis), jakožto přepojovací jádro zrakové dráhy.
- Specifická nesenzorická – „ motorická“ -jádra ventrální skupiny – nc. ventralis anterior, nc. ventralis medialis, nc. ventralis lateralis – aferentace jdou hlavně z mozečku, substantia nigra a bazálních ganglií. Eferentace míří k motorickým a premotorickým oblastem mozkové kůry. Slouží k převodu vzruchů z CRBL a basálních ganglií do kůry, jsou zapojena v přípravných motorických okruzích. Nuclei mediales et anteriores jsou zapojeny do okruhů limbického systému.
- Asociační – propojení specifických jader hrbolu – thalamu s asociačními oblastmi kůry, integrují senzitivní a senzorické podněty:
- Jádra nc. anterodorsalis, nc. anteroventralis, nc. anteromedialis, nc. lateralis dorsalis přijímají aferentace ze spodiny mezimozku a eferentace projektují do limbické kůry. Podílejí se na vzniku emocí, motivaci, pudů a drivu (úsilí, vervy, elánu).
- Nucleus lateralis posterior (aferentace z ncc. ventrales posteriores, eferentace vysílají vlákna do Brodmannových oblastí 5 a 6 mozkové kůry) a nucleus lateralis dorsalis (jehož mediální velkobuněčná část má oboustranné reciproční spoje s amygdalou, čichovými centry, laterálním hypothalamem a orbitotemporální kůrou, kdežto jeho laterální malobuněčná část má reciproční spojení s Brodmannovými oblastmi 9, 10, 11 a 12 prefrontální mozkové kůry).
- Nespecifické – ncl. reticularis thalami přijímá aferentace z retikulární formace kmene a z mozkové kůry a své eferentace směřuje k ostatním thalamickým jádrům a do retikulární formace středního mozku. Jedná se o difuzní projekci do širokých korových oblastí a basálních ganglií, převádí hlavně vlivy síťové formace (FR).
Dělení dle způsobu zapojení:
- Relé (přepřahací, přepínací) jádra – mají jeden hlavní podkorový vstup a projikují do přesně vymezené korové oblasti.
- integrační jádra – přijímají signály z více podkorových vstupů, projikují do několika asociačních oblastí kůry.
Stimulováním receptorů dané modality nebo elektrickou stimulací specifických senzorických jader talamu s frekvencí 8–15/s dojde k vyvolání tzv. augmentační reakce (augmenting response – narůstající odpověď) v odpovídajících projekčních oblastech mozkové kůry.13
13 Wikipedie: Otevřená encyklopedie: Thalamus [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Thalamus&oldid=11440155
Naopak elektrická stimulace talamických jader s difůzní projekcí vyvolá odpověď v celé mozkové kůře, která se liší od frekvence stimulace. Jedná se buď o desynchronizační odpověď (při frekvenci 100–300/s), nebo tzv. náborovou reakci (recruiting response, při frekvenci 6–10/s).
Thalamus je zapojen jako přepojovací stanice všech senzitivních drah mimo čichových. I z čichových drah však přijímá kolaterální vlákna. Dráhy vstupují cestou lemniscus medialis, lemniscus lateralis, lemniscus trigeminalis, tractus spinothalamicus a trigeminothalamicus, brachium colliculi inferioris et superioris. Thalamická jádra (mimo retikulárních) vysílají axony do specifických oblastí mozkové kůry a z kůry také vlákna přijímají. Vjemy přijímané thalamickými jádry jsou vždy sdíleny i mozkovou kůrou, thalamus a mozková kůra se navzájem ovlivňují. Thalamus představuje také důležitou přepojovací strukturu v senzoricko – motorických okruzích nezbytných pro volní pohyb.
- Lamina medullaris medialis – vnitřní ploténka bílé hmoty – je složena z vláken propojující jádra thalamu navzájem. Obsahuje intralaminární jádra. Rostrálním směrem se vidlicovitě rozděluje.
- Lamina medullaris lateralis - zevní ploténka bílé hmoty mezi retikulárními a laterálními jádry thalamu.
- Radiatio anterior thalami - svazky myelinizovaných vláken oboustranně spojující ncll. anteriores thalami s gyrus cinguli. Běží v crus anterior capsula interna.
- Radiatio centralis thalami - vějířovitě uspořádané svazky v crus posterior capsulae internae. Spojují především ncl. ventralis posteromedialis a posterolateralis s parietální kůrou senzitivní korové oblasti.
- Radiatio posterior thalami – vlákna běží v retrolentikulární části capsula interna. Spojují thalamus s occipitálním lalokem koncového mozku.
- Pedunculus thalamicus inferior - svazek vláken spojujících thalamus s hypothalamem.
- Fibrae intrathalamicae - spojení jednotlivých jader thalamu navzájem.
- Radiatio optica (Gratioletův svazek) - část zrakové dráhy. Probíhá z corpus geniculatum laterale v crus posterior capsulae internae, kolem týlního rohu postranní komory do primární zrakové kůry.
- Radiatio acustica – část sluchové dráhy, spojuje corpus geniculatum mediale s gyri temporales transversi. Probíhá v retrolenticulární části crus posterior capsulae internae.
- Fasciculus thalamicus – leží pod thalamem, nad zona incerta, na řezu ve Forelově poli H1. Obsahuje ansa lenticularis, fasciculus lenticularis (oba svazky vedou vlákna z globus pallidus do ncll. ventrales thalami) a vlákna z mozečku do thalamu.
- Ansa lenticularis – svazek vláken z globus pallidus do nuclei ventrales thalami, kolem předního okraje capsula interna.
- Fasciculus lenticularis – svazek vláken z globus pallidus, do nuclei ventrales thalami, skrze capsula interna. Probíhá přes Forelovo pole H2.
- Ansa peduncularis – spoje mezi thalamem a claustrem.
- V thalamu dochází k přepojování aferentací, které dále směřují do mozkové kůry – senzitivní, zrakové, sluchové a chuťové. Při poruchách thalamu dojde ke snížení prahu pro bolest, taková bolest se pak označuje jako thalamická a není dobře lékově zvládnutelná.
- Činnost thalamu je podřízena mozkové kůře. Kortikothalamické a thalamokortikální spoje ovlivňují bdělý stav člověka.
- Stimuly, které do thalamu míří z hypothalamu a zpětně i z limbické kůry, způsobí, že se thalamus podílí na některých vegetativních funkcích jako je zblednutí nebo zčervenání v obličeji, změny ve frekvenci tepu, dále na změnách nálady – veselosti, smutku, zlosti nebo rozmrzelosti. Tyto děje jsou v dospělosti tlumeny mozkovou kůrou, ale v plném rozsahu se projevují v dětském věku, kdy útlum není vyvinut nebo u lidí s poruchou funkce mozkové kůry.
- Thalamus se podílí také na ovlivnění stoje a chůze, prostřednictvím převodu vzruchů z mozečku do mozkové kůry. Při jeho poruše může vzniknout mírná mozečková ataxie.
Pozn.:
Nuclei reticulares thalami - jádra uložená zevně od lamina medullaris externa, navnitř od capsula interna. Představují nejrostrálnější oddíl retikulární formace. Afferentní vlákna dostávají z kůry koncového mozku a retikulární formace. Výstupní vlákna směřují hlavně do ostatních skupin thalamických jader. Jejich funkce není zcela objasněná, ale podle zapojení mohou tato jádra být prostředníkem vlivu kůry na činnost thalamu.
↓ rozbalit celou poznámku ↓Pozn.:
Nuclei reticulares thalami - jádra uložená zevně od lamina medullaris externa, navnitř od capsula interna. Představují nejrostrálnější oddíl retikulární formace. Afferentní vlákna dostávají z kůry koncového mozku a retikulární formace. Výstupní vlákna směřují hlavně do ostatních skupin thalamických jader. Jejich funkce není zcela objasněná, ale podle zapojení mohou tato jádra být prostředníkem vlivu kůry na činnost thalamu.
Nuclei intralaminares thalami - skupina drobných jader uložených uvnitř lamina medullaris interna. Největší z nich je ncl. centromedianus. Jádra přijímají vlákna retikulární formace, tr. spinothalamicus a tr. trigeminothalamicus. Efferentní vlákna směřují k ostatním thalamickým jádrům (dále po přepojení do kůry koncového mozku) a ke corpus striatum. U těchto jader se předpokládá vliv na uroveň vědomí a bdělosti jedince.
Nuclei mediani thalami - jádra uložená pod ependymem třetí komory a v adhesio interthalamica. Jsou obousměrně propojena s hypothalamem. Převádí vzruchy z retikulární formace do frontální a limbické kůry.
Nuclei anteriores thalami - jádra uložená v rozštěpu lamina medullaris interna. Podmiňují tuberculum anterior thalami. Přijímají tractus mammilothalamicus, vlákna z gyrus cinguli a hypothalamu. Jsou zapojena do Papezova okruhu limbického systému (emoční ladění, paměť). Patří sem ncll. anterodorsalis, anteromedialis, anteroventralis).
Nuclei mediales thalami - skupina jader mezi ncll. mediani a lamina medullaris interna. Největším je ncl. mediodorsalis. Toto jádro má oboustranné propojení s frontální kůrou, hypothalamickými jádry, přijímá informace z ncl. amygdaloideum a retikulární formace. Jádra jsou odpovědná za integraci senzitivních informací somatických i viscerálních a za vztah těchto informací k emočnímu ladění a subjektivnímu stavu.
Nuclei dorsales thalami (laterales) - jádra uložená v dorzální řadě mezi lamina medullaris interna a externa. Jsou to zepředu dozadu ncl. lateralis dorsalis a ncl. lateralis posterior. Zapojení těchto jader není zcela známo, mají spoje s parietální a limbickou kůrou a ostatními jádry thalamu.
Nuclei ventrales laterales thalami (ventrolaterales) - jádra uložená ve ventrální řadě mezi lamina medullaris interna a externa. Zepředu dozadu – ncl. ventralis anterior, ncl. ventralis lateralis, ncl. ventralis posterior. Nucleus ventralis posterior je rozdělen do ncl. ventralis posteromedialis a ncl. ventralis posterolateralis.
Nucleus ventralis anterior – jedno z jader ventrolaterálních. Má spoje s retikulární formací, substantia nigra, corpus striatum a premotorickou kůrou. Předpokládá se vliv jádra na aktivitu motorické kůry.
Nucleus ventralis lateralis - jedno z jader ventrolaterálních. Zapojené také do motorických okuhů. Hlavní aferentace je z mozečku, ale také přepojuje vlákna přicházející z ncl. ruber. Eferentní dráhy směřují do motorické a premotorické kůry. Pravděpodobně také ovlivňuje aktivitu motorické kůry.
Nucleus ventralis posteromedialis - jedno z jader ventrolaterálních. Přijímá senzitivní informace z oblasti hlavy (včetně chuťových) cestou lemniscus trigeminalis a tr. trigeminothalamicus. Eferentní spoje běží do primárních senzitivních oblastí kůry v gyrus postcentralis.
Nucleus ventralis posterolateralis - jedno z jader ventrolaterálních. Přijímá senzitivní vjemy z trupu a končetin cestou tr. spinothalamicus a lemniscus medialis. Eferentní spoje běží do primárních senzitivních oblastí kůry v gyrus postcentralis.
Nuclei posteriores thalami - asociační jádra pod pulvinar thalami. Koordinují zrakové a sluchové informace. Přijímají vlákna z míchy, corpora geniculata a tecta mesencephala. Eferentní dráhy pokračují do asociačních oblastí kůry. Na tomto schématu nejsou znázorněna, nachází se za myšlenou úrovní řezu.
Nucleus corporis geniculati medialis - součást sluchové dráhy. Afferentní vlákna z colliculus inferior vytváří brachium colliculi inferioris. Corpus geniculatum přijímá vjemy z obou uší, ale hlavně z druhé strany. Efferentní vlákna směřují jako radiatio acustica do primární korové sluchové oblasti v gyrus temporalis superior. Na tomto schématu není znázorněno, nachází se za myšlenou úrovní řezu.
Nucleus corporis geniculati lateralis - součást zrakové dráhy. Je utvářeno šesti vrstvičkami šedé hmoty. Přijímá témeř všechna vlákna tractus opticus (mimo vláken, která směřují do area preoptica). V důsledku křížení vláken (axony gangliových buněk sítnice) v chiasma opticum, zpracovává corpus geniculatum laterale informace z temporální části sítnice stejnostranného oka a nasální části sítnice druhostranného oka. Efferentní vlákna vytváří radiatio optica, směřující k zrakové kůře v týlním laloku koncového mozku. Na tomto schématu není znázorněno, nachází se za myšlenou úrovní řezu.
↑ sbalit poznámku ↑ -
Podhrbolí – hypothalamus Obrázek č. 44 Obrázek č. 45
Funkčně je hypothalamus hlavním centrem pro řízení orgánů těla, reguluje mnoho aktivit tělesných orgánů. Mezi jeho funkce patří:
- Kontrola autonomního nervového systému - reguluje srdeční rytmus,tlak krve, pohyby trávící trubice, vylučování potních a slinných žláz a další orgánové aktivity.
- Kontrola emocionální odpovědi - oblati pro potěšení,vztek,sexuální energii a strach.
- Regulace tělesné teploty - jedná se o tělesný termostat – některé buňky podhrbolí jsou citlivé na teplotu krve a tím spouští chladící nebo zahřívací mechanismy dle potřeby- pocení, chvění. Centra v podhrbolí navozují horečku.
- Regulace pocitu hladu a žízně - dáno vnímáním koncentrace živin – hlavně cukru, a solí v krvi. Jádra pro iniciaci jezení se nachází v postranní části podhrbolí, jádra potlačující jezení se nachází v přední a vnitřní části.
- Kontrola chování - ovládání naší motivace pro příjem potravy, určuje objem snězeného jídla, ovlivňuje sexuální chování.
- Regulace cyklu spánek-bdění - reguluje složitý jev spánku, je zopdovědnýý za načasování spánkového cyklu.Jádra angažující se v této regulaci jsou jádra nadzraková – ncll. supraoptici a blízko uložené předzrakové jádra – ncl. praeopticus. Nadzrakové jádro funguje jako biologické hodiny, které regulují časování mnoha denních rytmů – cirkadiální rytmy. Přijímají informace z očí o světle či tmě cestou optického nervu a pak vysílají signály do předzrakového jádra. Odpovědí je spuštění spánku.předzrakovým jádrem. Jádra v blízkosti bradavkového tělíska způsobují probuzení. Na tomto časování se spolupodílí i šišinka.
- Kontrola endokrinního systému - ovládá sekreci hormonů podvěsku mozkového a ten reguluje další žlázy s vnitřní sekrecí – jedná se o hierarchický kaskádový systém se zpětnými vazbami – a tím mnoho dalších funkcí v těle.
- Tvorby paměti - jádra bradavkových tělísek jsou propojena s paměťovým systémem v mozku.
Podhrbolí – hypothalamus je součást mezimozku – diencephala pod mezimozkovým hrbolem – thalamem. Je rozložen od zrakového zkřížení – chiasma opticum po dolní okraj bradavkového tělesa – corpora mamillaria. Tvoří dno a dolní část laterálního ohraničení třetí mozkové komory. Kaudálně podhrbolí – hypothalamus navazuje na středomozkovou pokrývku – tegmentum mesencephali. Těsně před podhrbolím – hypothalamem je uložen předzrakový okrsek – area preoptica, funkčně s ním spojená. Vývojově hypothalamus odpovídá visceromotorické ploténce mezimozku – diencephala. Jeho význam spočívá v kontrole homeostázy organismu cestou řízení autonomního nervového a endokrinního systému. V souvislosti s autonomním nervovým systémem řídí přední hypothalamus parasympatikus a zadní hypothalamus sympatikus. Šedá hmota je složena z neuronů uspořádaných do dobře ohraničenách jader a nepřesně vymezených oblastí – areae. Podhrbolí – hypothalamus můžeme rozdělit na:
- přístřední -mediální a
- boční – laterální část,
podle hranice, kterou tvoří část krytová klenby – pars tecta fornicis probíhající do bradavkových těles – corpora mamillaria. Některá jádra zasahují do obou zón. Zepředu dozadu je možné mediální hypothalamus rozdělit na tři oblasti:
- zkříženou (nadzrakovou) – chiasmatickou (supraoptickou),
- hrbolovou (nálevkovohrbolovou) – tuberální (infundibulotuberální) a
- zadní (bradavkovou) – (mamilární).
Pro přehlednost uvádíme pouze některá (důležitější) jádra.
1. Oblast podhrbolová přední – regio hypothalamica anterior (zřížená) (chiasmatická část) zahrnuje krom dalších zejména: jádro příkomorové – nucleus paraventricularis, jádro nad zkřížením – ncl. suprachiasmaticus, jádronadzrakové – ncl. supraopticus, jádro podhrbolové přední – ncl. hypothalamicus anterior.
- Jádro příkomorové podhrbolí – nucleus paraventricularis hypothalami je jádro v blízkosti žlábku podhrbolového – sulcus hypothalamicus, obsahuje skupiny neuronů malo- i velkobuněčných. Produkuje oxytocin a vazopresin, které jsou axonálním transportem (pásmo podhrbolo-podvěskové – tractus hypothalamo-hypophysialis) předávány do zadního laloku hypofýzy.
- Jádro nad zkřížením – nucleus suprachiasmaticus - dobře ohraničené jádro. Aferentní vlákna přichází ze sítnice, jader bočního kolínkového tělíska – ncl. corporis geniculi lateralis, eferentní vlákna spojují jádro s ostatními jádry hypothalamu. Jádro nad zkřížením – suprachiasmatické jádro je zapojeno do řízení cirkadiálních rytmů.
- Jádro nadzrakové – nucleus supraopticus - jádro uložené nad zrakovým zkřížením – chiasma opticum. Produkuje vazopresin, v menší míře oxytocin. Hormony jsou vedeny axonálním transportem (pásmo podhrbolopodvěskové – tractus hypothalamo-hypophysialis) do zadního laloku hypofýzy.
- Přední jádro podhrbolí – nucleus anterior hypothalami - jádro leží za preoptickými jádry, je zapojeno do řízení parasympatického nervového systému.
- Jádra předzraková boční a přístřední – nuclei preoptici lateralis et medialis – skupina jader pod přední spojkou – commissura anterior, při koncové ploténce – lamina terminalis. Jsou zapojena do řízení parasympatického nervového systému a reprodukčního chování.
2. Střední podhrbolí – střední hypothalamus – políčko prostřední podhrbolové – area hypothalamica intermedia – krajina prostřední podhrbolová – regio hypothalamica intermedia (hrbolové podhrbolí) (tuberální hypothalamus) obsahuje mimo jiné: břišní -přístřední jádro podhrbolí – ncl. hypothalamicus ventromedialis, hřbetní-přístřední jádro podhrbolí – ncl. hypothalamicus dorsomedialis, jádra hrbolová – ncll. tuberales, jádro nálevkové – ncl. infundibularis.
- Hřbetní-přístřední jádro podhrbolí – nucleus dorsomedialis – jádro patří k hrbolovým-tuberálním jádrům, je uloženo nad předopřístředním-ventromediálním jádrem, kontroluje produkci hormonů předního laloku hypofýzy.
- Břišní-přístřední jádro podhrbolí – nucleus ventromedialis hypothalami – jádro patří k hrbolovým – tuberálním jádrům. Je uloženo nad nálevkou – infundibulem. Kontroluje produkci hormonů předního laloku hypofýzy a je zapojeno do řízení příjmu potravy (centrum sytosti).
- Jádra hrbolová – nuclei tuberales – malá jádra ve stěně nálevky – infundibula, uvolňují liberiny a statiny – působky ovlivňující tvorbu hormonů adenohypofýzy.
- Jádro obloukové (nálevkové) – nucleus arcuatus (infundibularis) – jádro v hrbolové-tuberální oblasti, těsně při stěně komory. Produkuje GHRH – releasing faktor růstového hormonu.
3. Zadní podhrbolí – zadní hypothalamus – políčko zadní podhrbolové – area hypothalamica posterior (bradavkové podhrbolí – mamillární hypothalamus) – zahrnuje jádra: jádro zadní podhrbolové – ncl. hypothalamicus posterior, jádra bradavkového tělesa – ncll. corpores mamillares, jádra hrbolobradavková – ncll. tuberomamillares.
- Jádro zadní podhrbolové – nucleus posterior hypothalami - nepřesně ohraničené jádro, které přechází do šedé hmoty v okolí středomozkového vodovodu – aquaeductus mesencephali.
- Jádra bradavkového tělesa prostřední a boční – nuclei corpores mamillares mediales et laterales - laterální jádro přijímá vlákna klenby-fornixu, mediální je zdrojem svazku bradavkvohrbolového – fasciculus mamillothalamicus.
4. Políčko boční předzrakové – area praeoptica lateralis – přední část laterálního hypothalamu, vývojově patří k telencephalonu.
5. Políčko boční podhrbolové – area hypothalamica lateralis - část laterálního podhrbolu -hypothalamu, zasahují sem některá jádra z prostředního podhrbolu – mediálního hypothalamu (jádro předzrakové – ncl. praeopticus, jádro nadzrakové – ncl. supraopticus, jádro boční hrbolové – ncl. tuberalis lateralis, jádro hrbolobradavkové – ncl. tuberomamillaris).
Jádro boční – nucleus lateralis – jádro boční podhrbolu – laterálního hypothalamu, je zapojené do kontroly příjmu potravy (centrum hladu) a tekutin
Hypothalamus pro zajištění své funkce – řízení řady nepostradatelných fyziologických funkcí – přijímá informace z těla prostřednictvím nervových spojení, složení krve a mozkomíšního moku. Hypothalamická jádra informace zpracovávají a stejnou cestou uplatňují svou kontrolní funkci. Mozkomíšní mok tak slouží také jako informační cesta mezi neurosekrečními buňkami hypothalamu a vzdálenými místy mozku. Nervovými drahami přijímá hypothalamus informace z následujících systémů: kolaterály ze somato- i viscerosenzitivních drah, kolaterály ze zrakové dráhy, spoje z čichového mozku – rhinencephala, kůry čelního -frontálního laloku, útvar mořského koníčka – hipokampální formace, mandle – amygdaly, dorzomediálních a středních jáder mezimozkového hrbolu – thalamu, středomozkové pokrývky – tegmenta mesencephali. Hlavní eferentní spoje pokračují do mozkového kmene a míchy pro praegangliové neurony autonomního nervového systému, přes RF (síťovou formaci) do předních jader mezimozkového hrbolu – ncll. anteriores thalami, do RF (síťové formace) středomozkové pokrývky – tegmenta mesencephala, do lemového- limbického systému. Podvěsek mozkový – hypofýza je s podhrbolím – hypothalamem spojena jednak portohypophysárním systémem cév a dráhou podhrbolo-podvěskovou – tractus hypothalamo-hypophysialis (viz hypophysis).
V přehledu uvádíme hlavní spoje podhrbolu – hypothalamu:
- Klenba – fornix – párový svazek vláken spojující hippokampovou formaci a bradavkové těleso – corpus mamillare. Obsahuje i vlákna spojující hippocampovou formaci pravé a levé strany, projekci hippocampu do septální oblasti telencephala, jader thalamu a hypothalamu. Fornixy jsou 2–3 mm silné, začínají ve spánkovém – temporálním laloku hemisféry jako klenbová ramínka – crura fornicis, spojení pravé a levé klebny – fornis dexter et sinister, vytváří klenbovou spojku – commissura fornicis. Pod mozkovým svalkem – corpus callosum pokračují klenbová těla – corpora fornicis a vpřed se rozbíhají v klenbové šíje – columnae fornicis.
- Vlákna hraničního proužku – fibrae striae terminales - obsahují vlákna obousměrně propojující mandlové jádro – amygdalu s jádry podhrbolí – hypothalamu. Svazek je uložený pod hraničním proužkem hrbolu – stria terminalis thalami a je součástí limbického (lemového) systému.
- Hřbetní podélný svazek – fasciculus longitudinalis dorsalis - propojuje nadřazená centra autonomního nervového systému v podhrbolí – hypothalamu s retikulární formací kmene a s visceromotorickými jádry hlavových a míšních nervů. Probíhá mozkovým kmenem.
- Hrbolový dřeňový proužek – stria medullaris thalami - soubor vláken při okraji mezimozkového hrbolu – thalamu pod hrbolobým proužkem – taenia thalami, směřující k uzdičce – habenule. Obsahuje vlákna z podhrbolí – hypothalamu, přepážky – septa, hippocampu, čichových oblastí do udičkových jader – ncll. habenullae. Je součástí limbického (lemového systému.
- Prostřední svazek koncového mozku – fasciculus medialis telencephali – z angličtiny MFB – medial forebrain bundle. Obsahuje vlákna z vývojově starých oblastí kůry a z RF do podhrbolí -hypothalamu. Součást limbického (lemového)systému.
- Hlavní bradavkový svazek – fasciculus mamillaris princeps – začíná z bradavkového těla – corpus mamillare, kde navazuje na klenbu fornixu a rozděluje se na bradavkokrytový svazek – fasciculus mamillotegmentalis a bradavkohrbolový svazek -fasciculus mamillothalamicus.
- Bradavkopokrývkový svazek – fasciculus mamillotegmentalis – vlákna jdou z bradavkového tělesa – corpus mamillare do retikulární formace středomozkové pokrývky – tegmenta mesencephala.
- Bradavkohrbolový svazek – fasciculus mamillothalamicus - vlákna jdou z tělesa bradavkového do předních hrbolobých jader – ncll. anteriores thalami.
- Bradavková stopka – pedunculus mamillaris - svazek vláken z tělesa bradavkového -corpus mamillare do retikulární formace kmene a odtud zpět do tělesa bradavkového -corpus mamillare.
-
Dolní hrbol, přední hrbol mezimozkový – subthalamus – corpus Luysi
Přední hrbol mezimozkový, dolní hrbol – subthalamus (thalamus ventralis, přední talamus) je jedním z oddílů mezimozku (diencephalon). Do této jeho části zasahují struktury středního mozku červené jádro – nucleus ruber a černá hmota - substantia nigra. Všechny útvary subthalamu mají nervové spoje a vztahy k hybnosti a jsou součástí mimopyramidového systému. (extrapyramidového).
Párová část diencephala je uložená mezi tegmentem mesencephala a thalamu, laterálně od hypothalamu. Laterálně je subthalamus ohraničen vlákny capsula interna. Vývojově odpovídá subthalamus somatomotorické zóně bazální ploténky diencephala. Některé struktury byly při vývoji odsunuty (černá hmota – síťová část -substancia nigra – pars reticularis, bledá koule – globus pallidus) do jiných částí mozku. Bílou hmotu subthalamu tvoří množství drah z míchy, kmene a mozečku do thalamu, dráhy spojující bazální ganglia s motorickými jádry thalamu.
Červené jádro – nucleus ruber má těsný vztah k regulaci svalového napětí a pohybu, stejně jako černá hmota - substantia nigra, která je nejdůležitějším zdrojem dopaminergních neuronů (produkujících dopamin).
Jádro předního hrbolu mezimozkového – podhrbolové jádro – nucleus subthalamicus (corpus Luysi) získává nervová vlákna z bledé koule - globus pallidus, odkud vychází i svazek čočkovitý - fascilus lenticularis, jehož vlákna končí ve Forelových polích H, H1 a H2. Podhrbolové jádro – nucleus subthalamicus je jádro tvaru bikonkávní čočky, uložené ventrolaterálně od zona incerta. Dorsálně jde svazek čočkovitý – fasciculus lenticularis (odděluje podhrbolové jádro – ncl. subthalamicus od nejisté zóny – zona incerta), bazálně je uloženo vnitřní pouzdro – capsula interna, cca 0,5 milion neuronů; hlavně projekčních, jen málo interneuronů, součást okruhů bazálních ganglií.
Vyskytuje se pouze u savců. Je zapojené do okruhů bazálních ganglií, patří k systémům řízení motoriky. Přijímá aferenty z boční bledé koule – globus pallidus lateralis a motorické korové oblasti, eferenty vydává do boční i přístřední bledé koule – globus pallidus lateralis i medialis. Poškození podhrbolového jádra – ncl. subthalamicus se projeví druhostranným hemibalismem.
Aferentní spoje: Obrázek č. 46
- Bledopodhrbolová – pallidosubthalamická vlákna tvoří svazek fasciculus subthalamicus mediátor: GABA → inhibiční vliv pallida na aktivitu neuronů
- Motorická, premotorická a doplňková motorická korová oblast (area 4,6) vlákna končí převážně v motorickém sektoru (dorsolaterální část jádra), mediátor: glutamát → excitační vliv mozkové kůry na aktivitu neuronů
- Meziploténková – intralaminární jádra, hrbolopodhrbolové – thalamosubthalamické spoje, excitační
- Černá hmota – substantia nigra (část hutná – pars compacta), dopaminergní projekce
- Švová – rafeální jádra retikulární formace, serotoninergní projekce
- Jádro stopkomostové – nucleus pedunculopontinus, cholinergní projekce
Eferentní spoje
- Bledá koule – globus pallidus (pallidum internum et externum), hlavní eferentní projekce, mediátor: glutamát → excitační vliv
- Černá hmota – substantia nigra
Nejistá zóna – zona incerta – proužek šedé hmoty pod mezimozkovým hrbolem -thalamem, laterálně přechází do jádra síťového hrbolu – nucleus reticularis thalami, jádro je uloženo v pokračování retikulárních jader thalamu. Pro obdobné zapojení jako retikulární formace je k ní řazena zona incerta.
spoje
aferentní spoje jdou do mozkového kmen, thalamus a mozková kůra
eferentní spoje:
- vzestupné – končí v hypothalamu, thalamu a mozkové kůře
- sestupné – končí v recipročních strukturách mozkového kmene
- většina vláken končící v thalamu a mozkové kůře jsou inhibiční (GABAergní)
Subthalamem prochází svazky vláken, které vystupují z mozkového kmene, mozečku a bazálních ganglií a míří do mezimozkového hrbolu – thalamu:- Hrbolový svazek – fasciculus thalamicus – dorzálně od nejisté zony – zona incerta, obsahuje bledohrbolová – pallidothalamická, mozečkohrbolová – cerebellothalamická vlákna, prostřední kličku – lemniscus medialis, míchohrbolovou dráhu – tractus spinothalamicus, na řezu patrný jako Forelovo pole H1 (bílá hmota mezi hrbolem -thalamem a nejistou zónou -zona incerta), obsahuje aferentní dráhy mezimozkového hrbolu – thalamu (vlákna bledohrbolová – pallidothalamická).
- Čočkovitý svazek – fasciculus lenticularis - vystupuje z mediálního segmentu bledé koule – globus pallidus (z jeho mediální části), ve Forelově poli H2 (bílá hmota mezi nejistou zónou – zona incerta a podhrbolovým jádrem – ncl. subthalamicus), obsahuje spoje bledohrbolové – pallido-thalamické, které beží přes vnitřní pouzdro – capsula interna.
- Smyčka čočkovitý – ansa lenticularis - vystupuje z mediálního segmentu bledé koule – globus pallidus (z jeho laterální části), zahrnuje vlákna bledohrbolová – pallido-thalamická, která probíhají kolem předního okraje vnitřního pouzdra – capsula interna.
- Prerubrální pole – na řezu vytváří Forelovo pole H (bílá hmota rostrálně od červeného jádra – ncl. ruber, mediálně od nejisté zóny – zona incerta). Zahrnuje vlákna z míchy, kmene a mozečku (přístřední klička – lemniscus medialis, trojklanná smyčka – lemniscus trigeminalis, míchohrbolová dráha – tr. spinothalamicus…) a čočkovitou smyčkou – ansa lenticularis. Po připojení vláken z H2 pole, pokračují společně do H1.
- Podhrbolový svazek – fasciculus subthalamicus – propojuje podhrbolové jádro – nucleus subthalamicus a bledou kouli – globus pallidus.
-
Nadhrbolí – epithalamus
zahrnuje fylogeneticky staré části diencephala, je uložen dorzomediálně od mezimozkového hrbolu – thalamu, skládá se z uzdičkových – habenulárních jader, uzdičková spojka – commissura habenularum, zadní spojka – commissura posterior a šišinky – glandula pinealis (epifýza). Vzniká z ependymových gliových buněk, vylučuje hormon melatonin, který ovlivňuje cirkadiánní cykly těla, signalizuje přípravu na noční spánkový cyklus. Ovlivňuje zrání a stárnutí.
Nadhrbolí tvoří:
- Prostřední a boční uzdičková jádra – ncll. habenulares medialis et lateralis jsou malá skupina neuronů pod uzdičkovými trojúhelníky – trigona habenularum. Přijímají vlákna z mandlového jádra – ncl. amygdaloideus a z hipokampální formace, cestou hrbolového dřeňového proužku – stria medullaris thalami. Některá vlákna hrbolového dřeňového proužku - stria medullaris thalami se kříží a vytváří uzdičkovou spojku – commissura habenularum. Eferentní vlákna z uzdičkových jader – ncll. habenulares pokračují do mezistopkového jádra – ncl. interpeduncularis (uzdičkomezistopková dráha – tr. habenulointerpeduncularis) a středomozkového krytu – tecta mesencephala, do mezimozkového hrbolu – thalamu a síťové formace – retikulární formace mozkového kmene. Jádra jsou považována za centrum koordinace čichových, viscerosenzitivních a somatosenzitivních vjemů.
- Udičková spojka – commissura habenularum (habenula) - obsahuje vlákna z hrbolového dřeňového proužku – stria medullaris thalami, která se kříží do druhostranných uzdičkových – habenulárních jader.
- Šišinka – corpus pineale; epifýza - drobný konický útvar připoutaný svou stopkou k dorzálnímu obvodu mezimozku – diencephala. Je rozdělena neúplnými septy z pojivové káně, na malé lalůčky. Histologicky obsahuje gliální buňky, pinealocyty a kalcifikovaný materiál (mozkový písek – acervulus cerebri) jehož množství se s věkem zvyšuje. Epifýza je v současnosti považována za endokrinní žlázu schopnou působit na činnost hypofýzy, Langerhansových ostrůvků, nadledvin, parathyroidních žláz a gonád. Působky epifýzy se dostávají k cílovým strukturám krevní cestou nebo cestou mozkomíšního moku. Produkuje melatonin, který inhibuje uvolňování gonadotropních hormonů. Sekreční aktivita epifýzy vykazuje cirkadiální rytmy, zvyšuje se za tmy.
- Zadní spojka (nadhrbolová) – commissura posterior (epithalamica) – příčný svazek vláken mezi ustím mezimozkového vodovodu – aquaeductus mesencephali a odstupem epifýzy. Obsahuje vlákna komisurální, spojující zadní jádra hrbolu – thalamu, horní hrbolky – colliculi superiores a jádra předkrytová – praetektální obou stran (vždy s odpovídající druhostrannou strukturu) a projekční, která převádí vlákna z vmezeřeného jádra – ncl. intersticialis a ncl. Darkševiči do prostředního podélného svazku – fasciculus longitudinalis medialis druhé strany.
- Předkrytové políčko – area pretectalis – oblast na hranici středního mozku -mesencephala a mezimozku – diencephala, rostrálně při horních (hlavových) hrbolků – colliculi superiores, obsahující překrytová jádra- ncll. pretectales. Je zde přepojovací centrum pupilárního reflexu.
- Hrbolový dřeňový proužek – stria medullaris thalami - soubor vláken při okraji mezimozkového hrbolu – thalamu, pod hrbolovým proužkem – taenia thalami, směřující k uzdičce -habenula. Obsahuje vlákna z podhrbolí – hypothalamus, přepážky – septum, hippocampu, čichových oblastí do uzdičkových jader – ncll. habenullae. Je součástí limbického systému.
- Uzdičkomezistopková (zahnutá) dráha – tractus habenulointerpeduncularis (retroflexus, Meynerti) - svazek vláken z uzdičkových jader – ncll. habenulares do jader mezistopkových – ncll. interpeduncularis. Přepojuje podněty z limbických a čichových oblastí mozkové kůry přes jádra mezistopková – ncl. interpeduncularis dále na RF kmene a somatomotorická a visceromotorická jádra kmene a míchy. Toto spojení je jednou z cest zprostředkujících reakce autonomního nervového systému na čichové vjemy a emoční podněty.
Pozn.: pubetas praecox, jet-lag disease – pásmová nemoc (anglicky jet lag, doslova „tryskáčové zpoždění“; také jet syndrome, doslova „tryskáčový syndrom“), v odborných publikacích někdy desynchronosis (desynchronóza), je únava a poruchy spánku plynoucí z narušení biorytmů po rychlém leteckém překonání několika časových pásem. Může způsobit i tzv. spánkovou obrnu nebo krátkodobou nespavost.Příznaky : únava, dezorientace, nevolnost, bolesti hlavy, nespavost, dehydratace a nechutenství, podrážděnost, iracionální chování. Aklimatizace může trvat několik dnů).18
18 Wikipedie: Otevřená encyklopedie: Pásmová nemoc [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=P%C3%A1smov%C3%A1_nemoc&oldid=10815161
↓ rozbalit celou poznámku ↓Pozn.: pubetas praecox, jet-lag disease – pásmová nemoc (anglicky jet lag, doslova „tryskáčové zpoždění“; také jet syndrome, doslova „tryskáčový syndrom“), v odborných publikacích někdy desynchronosis (desynchronóza), je únava a poruchy spánku plynoucí z narušení biorytmů po rychlém leteckém překonání několika časových pásem. Může způsobit i tzv. spánkovou obrnu nebo krátkodobou nespavost.Příznaky : únava, dezorientace, nevolnost, bolesti hlavy, nespavost, dehydratace a nechutenství, podrážděnost, iracionální chování. Aklimatizace může trvat několik dnů).18
18 Wikipedie: Otevřená encyklopedie: Pásmová nemoc [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=P%C3%A1smov%C3%A1_nemoc&oldid=10815161
Je to endokrinní žláza produkující hormony melatonin a pravděpodobně i trofický hormon (glomerulotrofin) a endogenní halucinogen DMT. V některých kulturách byla šišinka označována jako "třetí oko" a považována za hlavní sídlo ducha. Na povrchu je kryta vazivovým pouzdrem, uvnitř je tvořena nemyelizovanými nervovými vlákny, neurogliovými buňkami a hlavními buňkami, zvanými pinealocyty, opředenými sítí krevních kapilár. S přibývajícím věkem se začínají objevovat kulovitá tělíska tvořená hydroxyapatitem a uhličitanem vápenatým, zvaná mozkový písek (acervulus cerebri).
Šišinka je hlavním místem tvorby spánkového hormonu melatoninu, který je z šišinky uvolňován do krevního oběhu. U savců a vyšších živočichů je melatonin důležitým hormonem ovlivňujícím cirkadiánní a cirkanuální biorytmy, spánek, bdění a činnost pohlavních žláz. Hladiny melatoninu jsou silně závislé na střídání světla a tmy. Objevuje se také v oblasti sítnice nebo zažívacího traktu. Je tvořen z aminokyseliny tryptofanu a jeho produkce je řízena tzv. cirkadiánními hodinami v oblasti hypothalamu. Světelné záření (především pak o vlnové délce 484 nm – což odpovídá světlemodré barvě) jeho tvorbu snižuje, tma naopak jeho produkci podporuje. V noci je hladina melatoninu v krvi výrazně zvýšená. Aktivita šišinky je ovlivněna hypothalamem, který zprostředkovává informace o tom, kolik denního světla dopadá na sítnici. Denní světlo a sluneční svit má tedy vliv i na náš spánek.1
1 Wikipedie: Otevřená encyklopedie: Šišinka [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=%C5%A0i%C5%A1inka&oldid=11709830
(cirkadianní = cirkadiánní rytmus je biologický rytmus s periodou o délce 20–28 hodin (lat. circa = přibližně, dies = den), cirkanuální = periodické změny fyziologických funkcí organismů, které probíhají během jednoho roku).
Biorytmus je nějaký děj, který se opakovaně – cyklicky – objevuje u živého organizmu s určitou pravidelností. Tato pravidelnost může mít jistou míru nepravidelnosti. Délka cyklu může být velmi rozdílná od hodnot řádově v hodinách až po roční cyklus (nazývaný cirkanuální). Příkladem biorytmu je délka cyklu přibližně jeden den. Jedná se o jeden z biorytmů. Biorytmy jsou např. periodická kolísání aktivity a bdělosti. Perioda biorytmů bývá přibližně denní, měsíční a roční. Zakladatelem tohoto pojmu je rumunský vědec Franz Halberg.2
2 Wikipedie: Otevřená encyklopedie: Biorytmus [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Biorytmus&oldid=8830408
Trofický hormon epifýzy stimuluje produkci aldosteronu v nadledvině.
Během dne je šišinka hlavním zdrojem serotoninu – hormonu našeho vnitřního štěstí. Je to rovněž jeden z neurotransmiterů. Právě na serotoninu záleží, jestli nervový signál dorazí do všech částí těla.
Legendy říkají, že kdysi dávno měl každý člověk třetí oko. Navíc bylo viditelné a spočívalo uprostřed obočí, u kořene nosu. Každý z nás má třetí oko, ale málokdo ho dokáže využívat. Časem však člověk začal duchovně upadat. Někdy však může dojít i k spontánnímu otevření třetího oka, což se mnoha lidem stalo po úrazu či otřesu mozku. Někteří vědci míní, že třetí oko (tedy naše mimořádné schopnosti,jogíny nazývané siddhis) je malá žláza uprostřed mozku velikosti fazolového zrna, epifýza.
V tradici Východu i Západu se epifýza odedávna spojovala s duší a představivostí. Řecký anatom Herofilus, který žil v 4. století před Kristem, tvrdil, že epifýza kontroluje průtok myšlenek. Učenci novověku předpokládali, že je sídlem obrazotvornosti a díky ní má naše duše a mysl vliv na fyzické tělo.
Pro dnešní vědce je epifýza nesmírně zajímavou žlázou. Je citlivá na světlo a denní dobu, což zřejmě souvisí s hormonem melatoninem, který žláza produkuje a který redukuje stav spánku a bdění. Epifýza ho produkuje pouze v noci, ve tmě. Je známé, že u lidí, kteří se nacházejí ve stavu tranzu, vytváří epifýza větší množství melatoninu.
Víme, že melatonin udělal v současném světě velkou kariéru. Pomáhá například pilotům a letuškám poradit si se změnami časových pásem. Melatonin prý má vliv i na stárnutí, proto je nazýván také hormonem mládí. Protože se jedná o přírodní hormon, nelze ho patentovat a většina farmaceutických firem má ve své nabídce melaninové tablety, které hltají lidé stále častěji. Projevily se však u nich paranormální schopnosti? Neprojevily! Je to nepříjemná zpráva pro ty, kteří by si chtěli jednoduše a bez námahy otevřít třetí oko.
Jak si však třetí oko otevřít? Jsou lidé, kterým se to zřejmě nepodaří nikdy. Jsou to především "tvrdí" materialisté, kteří se duchovním věcem posmívají a tvrdí o nich, že jsou pouhým výmyslem. Otevřít třetí oko se hned nepodaří ani těm, kteří ho chtějí využít ke čtení cizích myšlenek či na podobné "aktivity". Pokud však ano, velmi rychle o tuto schopnost přijdou.
Aktivizování třetího oka má sloužit především pro vlastní duchovní vývoj, k tomu, abychom si uměli lépe poradit se životem, správně se rozhodovat, rozlišovat, rozvíjet svoji intuici. Metod na otevření třetího oka je několik. Většina z nich pochází od dávných jogínů a mudrců Východu. Jogíni většinou začínají dýcháním, které postupně prohlubují. Pro začátečníky je lepší naučit se ho pod vedením zkušeného učitele. Začátečníci si mohou pohodlně sednou v pozici faraona nebo do lotosového květu, ruce položit na kolena (dlaně směrem nahoru), palec a ukazovák spojit. Začneme pomalu a hluboce dýchat bránicí. Nádech (počítáme jedna, dva, tři, čtyři), výdech (jedna, dvě, tři, čtyři). Párkrát se důkladně nadechneme a vydechneme, přičemž můžeme pocítit lehkou závrať hlavy – tak působí zvýšená dávka kyslíku. Takovým způsobem se rychleji zbavíme dotíravých myšlenek. Koncentrujeme se na dech a potom se v mysli soustředíme na bod mezi očima. Můžeme se na sebe dívat i v zrcadle – tehdy si uprostřed obočí přilepíme modré kolečko nebo kamínek a představujeme si, že na tomto místě vzniká energetický vír, zářící modrou nebo fialovou barvou. Nečekejme, že se nám třetí oko otevře okamžitě. Musí uběhnout týdny i měsíce každodenního cvičení dlouhého nejméně 30 minut denně.
Po staletí si okultisté a spirituální mistři mysleli, že tato žláza je "sídlem duše" – což je výraz, který zpopularizoval Descartes (1662).
Descartes byl posedlý pochopením, kdo jsme. Zpochybňoval vše – dokonce i názor, že se můžeme poznat. Zjistil, že smysly lze oklamat, že většina toho, co si myslíme, že známe, je skutečně iluze, a nakonec bojoval s možností, že naše vlastní identita coby jedinců není rovněž skutečná. Ale nakonec došel k závěru, že pokud je možné pochybovat o naši vlastní existenci, tak musí existovat "něco", co je schopné tuto pochybnost zažít. A toto něco je naše skutečná já. Jeho slavný výrok se dochoval: "Cogno ergo sum – myslím, tedy jsem".
"Ačkoliv duše je spojena s celým tělem, existuje tělesná část (šišinka), ve které plní svoji funkci více, než kde jinde… Šišinka je mezi průchody obsahující zvířecí duchy (vůdčím důvodem a nosičem pohybu a vjemu) tak ponořená, že jimi může být hnána…; a tento pohyb přenáší na duši… Pak opačně, tělesný stroj je sestaven tak, že kdykoliv žlázou tam či onam pohne duše, nebo jakákoliv jiná příčina, tlačí na zvířecí duchy, které ji obklopují do pórů mozku." – Descartes
Dnes, s porozuměním počítačům, můžeme s Descartem souhlasit. Že proces "myšlení" může být součástí neuronových obvodů, synapsí a neurotransmiterů, které existují v našem hmotném mozku. Skutečné já je mnohem hlouběji. Používá proces myšlení, ale zůstává mimo něj, proces pozoruje. Skutečná já je v podstatě samotné vědomí.
Tyto dva fenomény jsou očividně spojené: vědomí má vzájemný vztah s procesem myšlení. Takže skutečnou otázkou je, jak toto rozhraní hmotné mysli a spirituálního vědomí funguje. Pokud dokážeme pochopit, jak vědomí interpretuje naše myslící já, pak snad můžeme procesy zvrátit a stát se vědomými vědomí – našeho skutečného já. To je osvícení.
Jako obvykle, při provádění výzkumu šišinky mozkové byla odhalena některá úžasná fakta. Nejen, že se zdá, že tato neskutečně malá žláza souvisí s aktivitou psí a paranormálními schopnostmi, ale je extrémně citlivá na prostředí způsoby, které odhalují náš zjevný materialismus a pandemickou depresi.
Šišinka mozková v historii
Ačkoliv je to běžně přisuzováno Descartovi, tak názor, že šišinka mozková je orgánem rozhraní mezi duchem člověka, který tam získává přístup, a oživeným lidským tělem, má původ u řeckého lékaře jménem Herophilus. Tři sta let před Kristem Herophilus pitval těla a dokumentoval, co vidí. Jeho specialitou byl reprodukční systém a mozek.
Před Herophilem lidé věřili, že "výkonným centrem" lidského vědomí je srdce. Egyptské mumie měly srdce pečlivě nabalzamované a zachované, zatímco mozek byl odstraněn přes nosní dírky a neobřadně vyhozen. Ale Herophilus věděl, že mozek je řídícím centrem a rozlišoval mezi různými částmi mozku a vyhodnocoval různá chování s nimi související.
Herophilus si povšiml, že malý šišinkový útvar je jen jeden, na rozdíl od dalších struktur mozku, které mají zrcadla v levé a pravé polokouli. Je to první žláza, která se utváří v plodu, a je rozpoznatelná již ve 3 týdnech. Je také silně živena. Šišinka mozková je zásobována nejlepší krví, kyslíkem a směsí živin, v lidské anatomii dostupných, hned po ledvinách (jejichž funkcí je filtrovat nečistoty z krve). Díky této jedinečné a zvláštní anatomické konfiguraci Herophilus správně došel k závěru, že hraje značnou roli ve vědomí a je branou do našeho skutečného já.
Herophilovi je připisován "vynález" vědecké metody, dogmatu každého dnešního výzkumu, tedy z postulování teorie a provedení experimentů, aby se potvrdila nebo vyvrátila. Je pak ironické, že vědeckému výzkumu šišinky mozkové se bylo pečlivě vyhýbáno, kvůli její spojitosti se "spirituálním" fenoménem – u kterého není možné empirické prověřování. Teprve nedávno se vědci začali zajímat o získání větších znalostí o této záhadné žláze.
Herophilus napsal o anatomii mnoho svazků a ilustroval je, stejně jako Leonardo da Vinci činil o mnoho století později, ale jeho práce bohužel nepřežila zničení velké knihovny v Alexandrii, kde byly uloženy.
Třetí oko
V r. 1884 oblast komparativní anatomie odhalila důkazy, že lidské orgány lze vystopovat až k nižším obratlovcům. F. Ahlborn zjistil, že šišinka mozková byla původně fotoreaktivním orgánem, u ryb a obojživelníků, a někdy se nacházela mimo lebku, těsně pod pokožkou. Její funkce coby spouštěče reprodukce byla zaregistrována zhruba o 70 let později. Šišinka mozková je zodpovědná za vyhodnocování délky dne a noci, výpočet správného období pro páření a nárůstu sexuálního puzení.
V r. 1958 Aaron Lerner objevil melatonin, klíčovou molekulu vyráběnou v šišince mozkové z dalšího běžného neutransmiteru serotoninu. Rovněž potvrdil fakt, že produkce melatoninu se liší, ustává za dne a zvyšuje se krátce po setmění. Melatonin, jak bylo zjištěno, je zodpovědný za naši relaxaci a usnutí.
Po jistou dobu se nevědělo, jak tato malá žláza, pohřbená hluboko uprostřed lidského mozku, může vnímat světlo nebo temnotu. Ale pak bylo objeveno, že existuje k šišince spojení od sítnice, která, což je dosti podivné, také obsahuje melatonin. Okamžitě byla šišinka mozková označena za "třetí oko", a kvůli tomu, že se nachází na jedné ze sedmi čaker, byla známá jako centrum spirituální a parapsychologické energie.
Když se v 60. letech staly populární halucinogeny, zájem o pozměněné stavy vědomí, coby prostředku ke spiritualitě, vyvrcholil. Výzkumníci objevili, že LSD se v šišince a hypofýze koncentruje. Pozměněný stav byl vytvořen schopností LSD napodobovat serotonin na synapsích (kde spolu komunikují neurony). Protože jak serotonin, tak melatonin jsou koncentrovány v šišince, toto "třetí oko" bylo považováno za bránu do vědomí – to, co autor Alduos Huxley nazval "dveře vnímání".
Dokonce i guru šišinku komentovali a přirovnávali ji k jedné z čaker:
"Všechny parapsychologické systémy mají své fyzické aspekty v těle… Když je adžna čakra fyzickým ekvivalentem šišinky mozkové, které dlouho mátla doktory a vědce ohledně své přesné funkce… Jogíni, kteří jsou vědci jemné mysli, vždy mluvili o telepatii jako o "siddhi", parapsychologické moci pro myšlenkovou komunikaci a jasnovidectví atd. Médiem takové siddhis je adžna čakra, a její fyzickým mezníkem je šišinka mozková, která je spojena s mozkem. Bylo uvedeno velkými jogíny… že šišinka mozková je receptor a vysílač jemných vibrací, které přenáší myšlenky a parapsychologické fenomény vesmírem." – Satyananda, 1972. Ale podporuje něco z toho Herophilova "vědecká metoda"?
Bylo provedeno mnoho experimentů s tak zvaným fenoménem psí (paranormálnem) v 70. letech. Dixon (1) zjistil, že šišinka mozková je v této aktivitě primární. Povšiml si, že aktivita psí je nejúspěšnější v uvolněném ospalém stavu – podobně jako u aktivity vyvolané melatoninem. Další studie zjistily korelaci s melatoninem a praktikováním jogínských meditací, které měly být údajně nezbytným předpokladem pro "osvícení".
"Jogínské praktiky prováděné 3 měsíce měly za následek zlepšení kardiorespiračního výkonu a psychologického profilu. Plazmový melatonin tak vykázal nárůst po třech měsících jogínských praktik. Systolický krevní tlak, diastolický krevní tlak, střední arteriální tlak a ortostatická tolerance nevykazovaly nějakou významnou korelaci s plazmovým melatoninem. Nicméně maximální hladina melatoninu v noci u jogínské skupiny vykázala značnou korelaci (r=0,71, p < 0,5) s bodováním pohody a zdraví.
Tato pozorování naznačují, že jogínské praktiky lze použít jako psychologicko-fyziologický stimul ke zvýšení endokrinní sekrece melatoninu, který je pak zodpovědný za zlepšené vnímání pohody."
Dramatičtější důkazy přichází z mystických zkušeností a "zkušeností blízké smrti", vyvolanými chemikálií zvanou dimetyltryptamin (DMT). Tradiční "symbol" jihoamerických šamanů, DMT, silně připomíná jak melatonin, tak serotonin, a vyskytuje se přirozeně v některých tropických rostlinách, jakož i v lidském těle. Panuje názor, že uložený DMT je uvolňován ze šišinky mozkové těsně před smrtí, což způsobuje mimotělesné zážitky a mystické vize, hlášené přeživšími NDE (prožitek blízké smrti, near-death experience). Zatímco nejsou žádné empirické důkazy spojující šišinku mozkovou s produkcí DMT, její souvislost s melatoninem (ze kterého je vyvářen) tuto možnost zřejmě silně podporuje.
"Protože jde o endogenní, či přirozeně se vyskytují lidské psychedelium, věřil jsem, že může zprostředkovat spontánní psychedelické zážitky, jako NDE a mystické stavy. Také jsem považoval šišinku mozkovou za pravděpodobný zdroj tohoto endogenního DMT; jako taková by mohla být šišinka "žlázou ducha". (Strassman, Rick, J.,2001)
Lékařský výzkumník, JC Callaway (1988), tvrdil, že přirozeně se vyskytující DMT může souviset s fenoménem vizuálních snů, kdy úroveň DMT v mozku je periodicky zvyšována, aby se vyvolaly vizuální obrazy a možná i další paranormální stavy mysli. Takže ačkoliv empirické důkazy ezoterických funkcí šišinky mozkové nebyly nalezeny, zdá se, že tato myšlenka stojí za prozkoumání.
Mozkový písek
Anatomové ukázali, že lidská šišinka mozková je nejaktivnější v mládí, těsně před pubertou. To by dávalo smysl, kdyby šišinka nějak řídila reprodukční systém, zapínala nezbytné hormony, které představují dospělost. Ale po pubertě šišinka mozková, zdá se, že vápenatí. Akumuluje to, čemu se říká "mozkový písek". Ačkoliv šišinka mozková vykazuje toto zvápenatění, nezdá se, že by mizela tvorba melatoninu. V podstatě se zdá, že zvápenatění nemá žádný významný dopad. Ale ukázalo se, že některé další faktory vážně brání fungování šišinky. To je důležité, protože nedávné objevy ukázaly, že melatonin je klíčový pro imunitní systém, kdy nižší hladina je spojována s rakovinou prostaty a prsu (4) – dvěma hlavními příčinami předčasné smrti. Rovněž reguluje metabolismus krevního cukru. Studie ukazují, že pracovníci nočních směn dostanou s větší pravděpodobností cukrovku II, než jejich denní kolegové. Sezónní afektová porucha (SAD) je také spojována s deficitem melatoninu a serotoninu, ačkoliv mechanismus není zcela znám. Ze všech těchto účinků špatného zdraví je možnost dosažení "osvícení" s nedostatkem melatoninu trochu vzdálená!
Útoky na šišinku z okolního prostředí
Zasahování do šišinky mozkové z prostředí je všudypřítomné. Dvěma největšími antagonisty jsou elektromagnetické záření a fluorid. Studie na křečcích ukázaly, že expozice poli o 60 Hz (přesně proud v elektrické síti) výrazně snížila schopnost šišinky mozkové vytvářet melatonin během nočních cyklů."…akutní expozice magnetickému poli opětovně ztlumila nárůst a potlačila trvání nočního nárůstu melatoninu." Tuto anomálii také ukázaly studie na lidech, kteří byli vystaveni magnetickým polím o 60 Hz. Je zneklidňující toto zjistit, protože jsme všichni doslova obklopeni elektrickými poli o 60 Hz, doma, v práci a venku, kde jsou transformátory, téměř na každé ulici. Před tímto typem záření není úniku, vyjma útěku do odlehlé neobydlené oblasti. Ale i tam by mohla být šišinka pod útokem. Vzpomínáte na HAARP? Ohřívač ionosféry vymyšlený a patentovaný Richardem Eastlundem, který byl rychle zabaven ministerstvem obrany, předán Raytheon a rozvinut do zbraňového systému? Část patentu popisuje, jak silné záření extrémně nízké frekvence (ELF) je schopné způsobit akutní otupělost a zasahovat do regulace tepla – fungování šišinky mozkové. Není pochyb, že tato náchylnost byla za ty desítky let od doby, kdy byla tato technologie ukradena, využita a vypilována a použita pro ďábelskou agendu. Nyní může existovat již globální mřížka "ohřívačů" umístěných tak, aby bylo možné ovládat populaci a bránit našemu spirituálnímu vývoji. Do 90. let nebyl proveden žádný výzkum, aby se zjistil dopad fluoridu na šišinku mozkovou. Fluorid je běžně (a někdy povinně) přidáván do téměř veškeré pitné vody, údajně aby se zabránilo zubním kazům. Jeho použití bylo vždy vysoce kontroverzní a výzkum vedlejších účinků je často znesnadňován. Melatonin, vytvořený v šišince mozkové, nejen pomáhá regulovat nástup puberty, ale pomáhá také chránit tělo před poškozením buněk způsobeným volnými radikály. Nyní je známo – díky pečlivému výzkumu dr. Jeniffer Luke z britské univerzity v Surrey – že šišinka mozková je primárním cílem akumulace fluoridu v lidském těle. Měkká tkáň dospělé šišinky mozkové obsahuje více fluoridu, než jakákoliv jiná tkáň v těle (hladina fluoridu -300 ppm), to je více, množství postačující k zastavení činnosti enzymů). Šišinka mozková také obsahuje tvrdou tkáň (krystaly hyroxypapatitu), a tato tvrdá tkáň akumuluje ještě více fluoridu (až 21,000 ppm), než jakákoliv jiná tvrdá tkáň v těle (dokonce než zuby, které má údajně chránit). Po zjištění, že šišinka mozková je cílem tolika fluoridu, provedla dr. Luke experimenty na zvířatech, aby zjistila, jestli může mít nahromaděný fluorid dopad na regulaci melatoninu. Zjistila, že zvířata ošetřovaná fluoridem měla nižší hladinu melatoninu v oběhu. Tato nižší hladina melatoninu byla doprovázena u fluoridem ošetřovaných samic dřívějším nástupem puberty.
Luke shrnula svá zjištění u lidí a zvířat následovně: "…lidská šišinka mozková obsahuje nejvyšší koncentraci fluoridu v těle. Fluorid je spojován s potlačenou syntézou šišinkového melatoninu u předpubertálních dívek a zrychleným nástupem sexuální dospělosti. Výsledky posilují hypotézu, že šišinka hraje roli v načasování nástupu puberty…" Doposud jsme odhalili dva hlavní útoky na normální fungování naší šišinky mozkové z prostředí. Pokud je to skutečně "sídlo duše", nebo "brána k osvícení", pak lidstvo úspěšně uteklo z této přirozené cesty sebepoznání. Prostředí, ve kterém právě nyní jste – sedíte před počítačem – má negativní účinek na vaši šišinku mozkovou. Takže pokládáme otázku: je něco, co můžeme udělat, abychom zlepšili zdraví tohoto důležitého útvaru v mozku, možná ano.
Naděje a šišinka
Rytmické dýchání a chvalozpěv
Šišinka mozková sedí na střeše třetí komory mozkové, přímo za kořenem nosu, a plave v malém jezírku mozkomíšní tekutiny. Nemá mozko-krevní bariéru jako mozkové struktury, kde jisté molekuly v obíhající krvi jsou blokovány. Místo toho se spoléhá na neustálou dodávku krve, dodávané prostřednictvím obzvláště bohaté vaskulární sítě.
Aktivity jako rytmické dýchání a chvalozpěvy vytváří na kyslík bohatou dodávku krve šišince mozkové. Mnoho jogínů popisuje, že cílem jejich dýchací techniky je zvýšit srdeční tep, a snížit krevní tlak. Ironicky se právě toto děje těsně předtím, než zemřeme. Je možné, že cílem této procedury je uvolnit DMT, o kterém mnozí věří, že je v šišince uložen. Pamatujte, DMT je psychedeliím tělu vlastní. Je to neurochemikálie specificky vytvořená k tomu, aby umožnila zaměřit naše myšlení dovnitř. Ačkoliv přesná chemická reakce není zatím chápána, DMT umožňuje, aby synaptické cesty unikly z běžných tras a umožnily se nové myšlenky a vjemy. Nadřízenost specifických hemisfér je zrušena, což umožňuje to, co lze popsat jako nesouměřitelné – faktické reality, které nemohou být racionálně (příčina-následek) vysvětleny, jsou však přesto skutečné. Všechny tyto nezbytné předpoklady jsou potřeba, abychom se postavili unikajícímu já.
Okamžitá karma
Zatímco nízkofrekvenční a magnetické záření snižuje produkci melatoninu, některé jiné typy záření, zdá se, šišinku mozkovou nabíjejí. V článku uveřejněném před mnoha lety na Viewzone, ESP a LST, bylo ukázáno, že psí, či paranormální schopnosti, jako dálkové pozorování či ESP, byly na vrcholu, když se střed galaxie Mléčná dráha nacházel na jistém místě na obloze. Přesným časem bylo 13:30 LST (místní siderický, hvězdný, čas. Jeden den hvězdného času – hvězdný den, siderický – netrvá 24 hodin, ale 23 h, 56 minut a 4,09 sekundy).
Jak ukazuje mapa galaxie, střed galaxie je bohatým zdrojem rádiových emisí. Toto záření by mohlo být "registrováno" šišinkou způsobem, který jsme zatím nepochopili. Fakt, že vrchol schopností nebyl v 12:00 LST, kdy je střed galaxie a maximální záření přímo nad hlavou, může být buď důsledkem určitého zpoždění v reakci šišinky, nebo může odrážet nějaký ideální úhel, který je pro dopad rádiových emisí na zemský povrch optimální.
Již bylo zjištěno, že šišinka mozková reaguje na světlo, rádiové frekvence a záření ELF – extrémy elektromagnetického spektra. Zvukové vlny jsou rovněž součástí tohoto spektra. Schopnost něčeho tak prostého, jako zvuku, pronikat naší lebkou a transformovat funkci a strukturu našeho mozku je těžko představitelná. Jihoameričtí a středoameričtí šamani používají ayahuascu, bylinný lektvar, který stimuluje DMT, pro psychologickou léčbu a spirituální zasvěcovací obřady.10
10 Zlaté Živé Světlo, Šišinka mozková - "sídlo duše" [online], Dostupné z WWW: http://www.duchovniprednasky.estranky.cz/clanky/krasne-texty-a-videa/sisinka-mozkova----sidlo-duse-.html
Dimethyltryptamin (DMT) je velmi silný, přírodně se vyskytující halucinogen produkovaný rostlinami a přítomný v stopovém množství i v lidském těle, tvoří se v šišince. Vykazuje podobnost s dalšími halucinogeny, LSD a psilocybinem. Chemicky je to bílá či nahnědlá aromatická krystalická látka.

Nejstarším užíváním DMT byl nález váčku obsahujícího šňupací směs yopo obsahující DMT, 5-MeO-DMT a bufotenin v pohřebišti v severním Chile. Poprvé bylo užívání domorodými obyvateli Karibiku zdokumentováno již při objevování Ameriky v roce 1496. V následujících staletích bylo zaznamenáno užívání yopo na různých místech v Jižní Americe. První záznamy o užívání ayahuascy, nápoje obsahujícího DMT a MAOI, v Amazonii pochází z poloviny 17. století, soudí se ale, že byla užívána již stovky, možná tisíce let předtím. Yopo i ayahuasca byly rituálními halucinogeny (Entheogeny), používanými tamějšími šamany například při náboženských obřadech, věštění či léčení apod. V Amazonii byly nalezeny i další entheogeny, ve kterých bylo identifikováno DMT, například oo-koo-hé. Informace, které o nich máme, jsou ale velmi sporé. Tyto tradice se na některých místech zachovaly dodnes.
Moderní vědou byl však DMT syntetizován až v roce 1931, jeho psychoaktivní účinky byly objeveny ještě o několik let později. Koncem šedesátých let se objevuje jako droga na černém trhu v USA, roku 1971 je v USA postaveno mimo zákon, což následuje i ve většině zemí světa. Na přelomu osmdesátých a devadesátých let zažívá DMT malý revival, například v Brazílii je legalizováno rituální užívání ayahuascy početnou církví Santo Daime i jinými církvemi. Neměli bychom také opomenout vliv Terence McKenny, známého popularizátora DMT té doby. V roce 1990 také poprvé po dvaceti letech začal výzkum DMT na lidech vedený Rickem Strassmanem.
DMT se minimálně ve stopovém množství vyskytuje pravděpodobně ve všech živých organismech. V některých rostlinách se ale vyskytuje v poměrně velké koncentraci (například Psychotria viridis, Mimosa hostilis nebo chrastice rákosovitá). V lidském těle je jeho přítomnost také prokázána a je vylučován pravděpodobně šišinkou, ze které se vylučuje chemicky příbuzný hormon melatonin. Jeho funkce v těle zatím nebyla uspokojivě objasněna, předpokládá se, že se mj. podílí na snění, zážitcích blízkých smrti (NDE), mystických prožitcích atd. DMT lze také vytvořit synteticky. DMT díky své extrémní síle není příliš populární drogou a od drogových dealerů není běžně k dostání, mnoho uživatelů si ho vyrábí pro vlastní potřebu.
DMT je extrémně silný, ač jen krátce účinkující halucinogen. Jedná se o tzv. agonistu serotoninu, mozkového neuropřenašeče, který řídí zpracovování informací, ale má vliv i na náladu a vnímání reality. Jelikož je DMT normálně přítomno v lidském těle (ač v malých množstvích), tělo je připraveno na jeho metabolizaci a působení
Zážitky uživatelů DMT jsou často velmi bizardní. Mezi časté fenomény patří například návštěva jiných dimenzí (hyperprostor) či komunikace s bytostmi je obývajícími, celkové radikální změny perspektivy, intenzivní kaleidoskopické vize i při otevřených očích, úplné opuštění vlastního těla a pocit umírání, výrazné změny ve vnímání času a rozměrů. Mezi méně časté pak změna barev (např. celý svět vypadá zlatě), zvukové halucinace, žaludeční nevolnost a další. Téměř všichni uživatelé ale tvrdí, že pro popis tohoto zážitku nestačí běžná slova a jakýkoliv pokus o popis je nutně velmi nepřesný a zkreslený. Tyto zážitky mohou být velmi živé (nezřídka jsou popisovány jako reálnější než každodenní realita) a v důsledku neočekávané intenzity pro mnoho lidí můžou být velice děsivé, někdy dokonce mohou způsobit přechodné psychické problémy. I zkušení uživatelé omamných látek říkají, že DMT je extrémně silný halucinogen a může vytvářet děsivé stavy.
Charakterově jsou také často přirovnávány k účinkům velkých dávek halucinogenních hub lysohlávek, jejichž hlavní aktivní látkou je psilocin, který se od DMT liší pouze jednou hydroxylovou skupinou.
DMT lze užívat orálně, ale pouze v kombinaci s MAOI, jinak není účinné. Při tomto způsobu užití účinky nastupují v řádu desítek minut a trvají několik hodin, jsou méně intenzivní a častěji jsou doprovázeny tělesnými účinky, jako je kupříkladu zvracení. Chemicky se tato kombinace vyskytuje v ayahuasce, rituálním halucinogenu amazonských šamanů, kteří ji využívají k léčení a komunikaci s duchy. Dále je možné DMT užít inhalačně (nejběžnější), injekčně nebo nasálně (šňupáním), případně i rektálně. V tomto případě účinky nastupují ihned a trvají 5–20 minut, ale jsou velmi intenzivní, intenzivnější než drtivá většina ostatních psychedelik v běžných dávkách. Někteří tvrdí, že právě tato látka umožňuje vnímaní věcí, které jsou normálně běžnému vnímaní nepřístupné.
DMT je v ČR na seznamu zakázaných substancí. Dávkování: Při inhalačním podání mohou být prahové účinky cítěny již od 2 mg. Běžná dávka se pohybuje okolo 30 mg a 50 mg je považováno za vysokou dávku. Mezi jednotlivci ale můžou být velké odchylky v dávce potřebné k dosažení srovnatelného účinku závisející na tělesné hmotnosti a psychických a fyzických předpokladech4
4 Wikipedie: Otevřená encyklopedie: Dimethyltryptamin [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Dimethyltryptamin&oldid=11474888
↑ sbalit poznámku ↑ -
Záhrbolí – metathalamus
součást mezimozkového hrbolu – thalamu zahrnuje kolínková tělíska – corpora geniculata – senzorická jádra zraková a sluchová.- Boční kolínkové tělísko – corpus geniculatum laterale - párový útvar spojený pomocí ramínka horního hrbolku – brachium colliculi superioris s horním (hlavovým) hrbolkem – colliculus superior (cranialis). Obsahuje jádro ve kterém se střídají vrstvy bílé a šedé hmoty jádro bočního kolínkového tělíska – ncl. corporis geniculati lateralis zapojené do zrakové dráhy.
- Přístřední kolínkové tělísko – corpus geniculatum mediale – párový útvar spojený pomocí ramínka dolního hrbolku – brachium colliculi inferioris s dolním (ocasním hrbolkem – colliculus inferior (caudalis). Obsahuje kompaktní jádro prostředního kolínkového tělíska – ncl. corporis geniculati medialis zapojené do sluchové dráhy.
-
Podvěsek mozkový – hypophysis, glandula pituitaria Obrázek č. 45
Podvěsek mozkový – hypophysis, glandula pituitaria je žláza s vnitřní sekrecí. Je uložena pod tvrdou plenou mozkovou – sedlovou přepážkou – diaphragma sellae – řasa tvrdé pleny mozkové, překrývající jámu podvěsku mozkového – fossa hypophysialis, či turecké sedlo – sella turcica, těla kosti klínové
Podvěsek mozkový je žláza s vnitřní sekrecí vyjímečná svou funkcí. Její hormony ovlivňují jiné žlázy s vnitřní sekrecí. Hypofýza je složená ze dvou částí, různého původu. Přední lalok adenohypofýza se vyvíjí z Ratkeho výchlipky ze základu nosohltanu. Zadní lalok neurohypofýza je vývojová součást mezimozku – diencephala.
Přední lalok hypofýzy – lobus anterior, žlaznatá část hypofýzy – adenohypophysis – je místem syntézy somatotropního hormonu (působí na tělesný růst a metabolismus, STH) a hormonů, které řídí jiné žlázy – adrenokortikotropní (stimuluje syntézu a vylučování hormonů kůry nadledvin, ACTH), thyreotropní (ovlivňuje činnost štítné žlázy,TSH), folikostimulační (u ženy stimuluje růst folikulů a produkci estrogenů, FSH), luteinizační hormon (u ženy působí na růst žlutého tělíska, ovulaci, produkci progesteronu, LH), prolaktin (ovlivňuje rozvoj a činnost mléčné žlázy). Hormony adenohypofýzy jsou pod vlivem regulačních hormonů – tzv. liberinů a statinů – produkovaných v jádrech hypothalamu. Do předního laloku hypofýzy jsou tyto dopravovány prostřednictvím hypofýzo-portálního oběhu.
Má 3 části:
- hrbolová část – pars tuberalis – část adenohypofýzy ve stopce hypofýzy
- vzdálenější část – pars distalis – největší část předního laloku hypofýzy, je tvořená sekrečními buňkami.
- prostřední část – pars intermedia – redukovaná část adenohypofýzy na hranici mezi adeno- a neurohypofýzou. Zaujímá asi 2 % objemu hypofýzy, produkuje melanostimulační hormon.
Zadní laloku hypofýzy – lobus posterior, nervová část hypofýzy – neurophyphysis, slouží jako depozitum hormonů uvolňovaných podhrobolovými -hypothalamickými jádry nadzrakovým a příkomorvým – ncl. supraopticus a ncl. paraventricularis. Tvorba hormonů hypothalamickými jádry se nazývá neurokrinie. Tyto hormony – vasopresin (antidiuretický hormon,ADH, adiuretin) a oxytocin – jsou do zadního laloku hypofýzy transportovány cestou podhrbolopodvěskové dráhy – tractus hypothalamohypophysialis, a jsou uvolňovány na povrchu fenestrovaných kapilár. Dalším produkovaným hormonem je antidiuretický hormon.
Pozn.: Antidiuretický hormon (ADH, též vazopresin) je peptidický hormon o délce 9 aminokyselin (takže se jedná o oligopeptid. V nejběžnější formě má sekvenci Cys-Tyr-Phe-Gln-Asn-Cys-Pro-Arg-Gly-amid (ale vyskytuje se i forma s lysinem na pozici 8 místo argininu). Název „antidiuretický hormon“ je odvozen ze skutečnosti, že působí proti diurézi, tedy tvorbě moči v ledvinách.5
5 Wikipedie: Otevřená encyklopedie: Antidiuretický hormon [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Antidiuretick%C3%BD_hormon&oldid=10879904
↓ rozbalit celou poznámku ↓Pozn.: Antidiuretický hormon (ADH, též vazopresin) je peptidický hormon o délce 9 aminokyselin (takže se jedná o oligopeptid. V nejběžnější formě má sekvenci Cys-Tyr-Phe-Gln-Asn-Cys-Pro-Arg-Gly-amid (ale vyskytuje se i forma s lysinem na pozici 8 místo argininu). Název „antidiuretický hormon“ je odvozen ze skutečnosti, že působí proti diurézi, tedy tvorbě moči v ledvinách. Synonymum „vazopresin“ odkazuje na mechanismus tohoto účinku – má schopnost zužovat periferní tepénky a vlásečnice v ledvinách. Vazopresin (a příbuzný oxytocin) se vyskytují zejména u obratlovců a zřejmě vznikly jistou genovou duplikací. Situaci však komplikuje skutečnost, že i u některých bezobratlých existují peptidy s podobnou strukturou a funkcí, jako vazopresin a oxytocin. ADH vylučují peptidergní neurony situované v hypothalamu (oblast mezimozku). Nevylučují však tento hormon do přilehlé tkáně, ale vysílají k tomu určené axony směrem do níže uložené neurohypofýzy – z ní pak probíhá vylučování do krevních kapilár. ADH se váže na (s G proteinem spřažené) vazopresinové receptory několika typů: V1 (V1a a V1b) a V2 receptory (V2). Známější je fyziologická role na V2 receptory: když se na tyto receptory ve stěně sběrných kanálků v ledvinách naváže ADH, dochází k zvýšenému průchodu vody z ledvin do krve a tedy k zahušťování moči. Z molekulárně biologického hlediska je toto zajištěno stimulací váčků s předem připravenými akvaporinovými kanály, a to zvýšením vnitrobuněčné koncentrace cAMP po navázání ADH na receptor V2 typu. Zajímavé jsou však i V1 receptory, na něž ADH rovněž působí. V1a se nachází na vnitřním povrchu některých cév, hlavně v močovém měchýři a v kůži, a je schopen zužovat tyto cévní systémy (ale fyziologická role tohoto je neznámá). V1b receptory se nachází na buňkách produkujících ACTH) v předním laloku hypofýzy a zdá se, že ADH má i roli v tělesné odpovědi na stres. U jiných obratlovců má vazopresin, jenž se váže na V1 receptory v mozku, i další funkce: ovlivňuje agresivní a naopak „přátelské“ chování, rozpoznání vlastních potomků a péče o potomstvo, ale zřejmě také např. reakci na kinetózu. Jedním z endokrinních projevů onemocnění zadního laloku hypofýzy (např. nádoru) je žíznivka (diabetes insipidus), při které je člověk neschopen zadržovat v organizmu vodu, tzn. člověk pak za den vylučuje 10 až 20 l vody – dechem, potem i močí. Ethanol tlumí tvorbu adiuretinu, a proto je po požití alkoholu vylučováno větší množství moči.5
5 Wikipedie: Otevřená encyklopedie: Antidiuretický hormon [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Antidiuretick%C3%BD_hormon&oldid=10879904

Oxytocin je peptidický hormon savců, který se syntetizuje v hypothalamickém jádru nucleus paraventricularis. Do krevního oběhu je uvolňován z neurohypofýzy. Uvolňuje se u žen po stimulaci prsu, dělohy a vagíny a v menší míře při doteku. Má vliv na porod i na kojení. Je možné, že u mužů zvyšuje kontrakce hladké svaloviny semenného provazce (vas deferens), které vystřikují sperma do močové trubice (urethry) a má vliv na budování vztahu mezi matkou a potomky (tzv. mateřská láska). Bývá také nazýván hormonem důvěry či lásky.6
6 Wikipedie: Otevřená encyklopedie: Oxytocin [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Oxytocin&oldid=11615045
- Způsobuje kontrakci myoepitelových buněk vývodů mléčné žlázy. Stah těchto buněk vede k ejekci mléka při kojení.
- Vede ke kontrakci hladké svaloviny dělohy, což při porodu zesiluje porodní činnost, v poporodním období usnadňuje vypuzení placenty a v průběhu šestinedělí napomáhá fyziologické involuci dělohy. Oxytocin je ve zvýšené míře secernován i při orgasmu, kdy pravděpodobně zahajuje děložní stahy usnadňující transport spermatu.
- V embryonální době napomáhá diferenciaci kardiomyocytů
- Podněcuje vznik příjemných pocitů a navozuje pečovatelské a ochranitelské chování u mužů i u žen (často nazýván jako hormon lásky).
- Zvyšuje empatii, kognitivní funkce, zvyšuje důvěru a snižuje strach, ovlivňuje sociální a sociálně sexuální chování, snižuje monotónní chování, zvyšuje emocionální projevy (ovlivňuje autismus), zvyšuje sexuální vzrušení, utváření vzájemných pout, zejména mezi pohlavími.
- Zvyšuje natriurésu – vylučování sodíku ledvinami, moduluje aktivitu hypotalamo-pituito-adrenální osy tím, že nepřímo blokuje adrenokortikotropní hormon a kortisol.

Hypofýzoportální systém – některá jádra hypothalamu (jádra popelavého hrbolu – ncll. tuberales, obloukové jádro – ncl. arcuatus, hřbetní a břišní přístřední jádro podhrbolí – ncl. dorsomedialis a ventromedialis) produkují liberiny a statiny, které aktivují nebo tlumí produkci hormonů předního laloku hypofýzy. Také tady mluvíme o neurokrinii. Hormony jsou axonálním prouděním (hrbolonálevková dráha – tractus tuberoinfundibularis) dopravovány do oblasti středové vyvýšeniny – eminentia mediana (drobná vyvýšeniny v nálevkovém výběžku – recessus infundibuli III. mozkové komory), kde je rozložena primární kapilární síť hypofyzárních tepen. Kapiláry se spojí do hypofyzárních portálních žil a ty se v předním laloku rozpadají na sekundární kapilární síť. Regulační hormony jsou krví přeneseny z oblasti primární kapilární sítě do adenohypofýzy, kde ovlivňují tvorbu a uvolňování svých cílových hormonů.7
7 Kikalová K., Machálek L., Holibka V., Kutal M., Bezděková M., Hypofýzoportální systém, NEUROANATOMIE I. - Struktury centrálního nervového systému [online], Univerzita Palackého v Olomouci, Dostupné z WWW: http://www.nan.upol.cz/neuro/cd557.html
Herringova tělíska jsou mikroskopicky pozorovatelné částečky neurosekretu.8
8 Kikalová K., Machálek L., Holibka V., Kutal M., Bezděková M., Herringova tělíska, NEUROANATOMIE I. - Struktury centrálního nervového systému [online], Univerzita Palackého v Olomouci, Dostupné z WWW: http://www.nan.upol.cz/neuro/cd563.html
Podhrbolopodvěsková dráha – tractus hypothalamohypophysialis – axony nadzrakového jádra – ncl. supraopticus a příkomorového jádra – ncl. paraventricularis, které transportují oxytocin a vasopresin do zadního laloku hypofýzy.9
9 Kikalová K., Machálek L., Holibka V., Kutal M., Bezděková M., Tractus hypothalamohypophysialis, NEUROANATOMIE I. - Struktury centrálního nervového systému [online], Univerzita Palackého v Olomouci, Dostupné z WWW: http://www.nan.upol.cz/neuro/cd564.html
↑ sbalit poznámku ↑Cévy zásobující podvěsek mozkový:
- Tepny: horní podvěsková tepna – arteria hypophysialis superior, dolní podvěsková tepna – arteria hypophysialis inferior – z karotického povodí.
- Žíly: podvěskové vrátnicové žíly – venae portales hypophysiales – žíly spojující primární a sekundární kapilární síť v adenohypofýze, žíla předního laloku – vena lobi anterioris.
| Obrázky ke kapitole: Mezimozek | |||
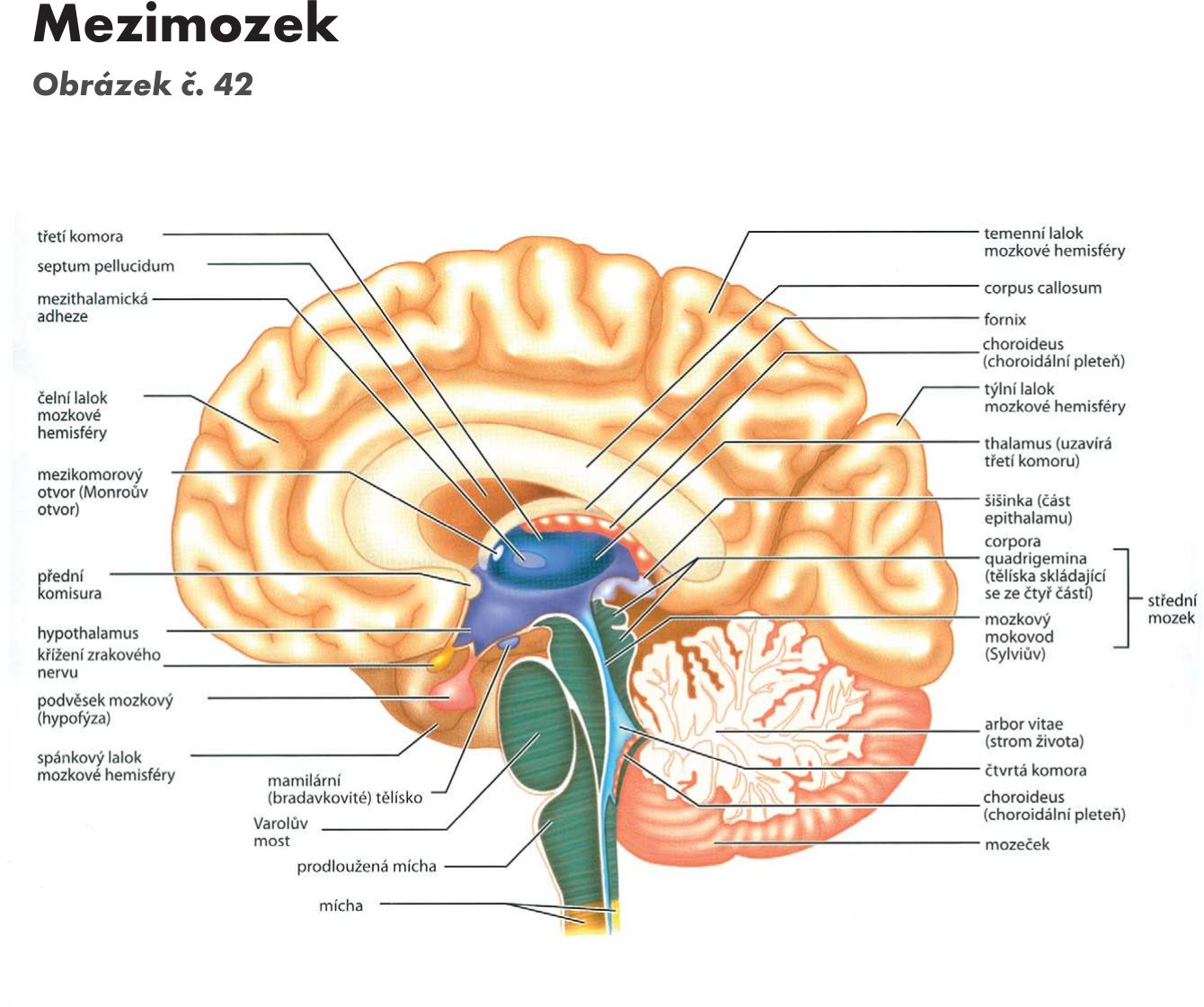
Obr. 42 Mezimozek
|
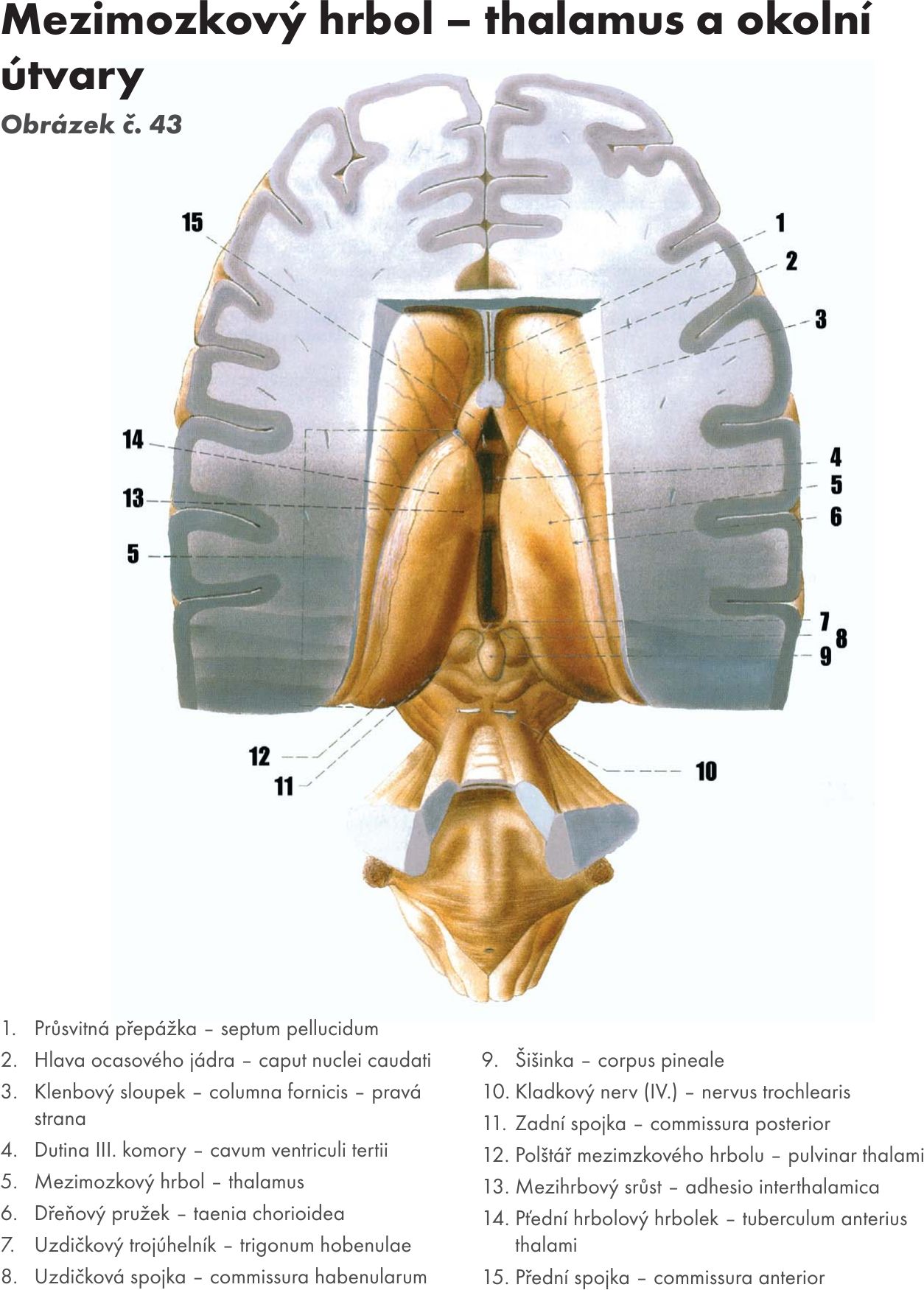
Obr. 43 Mezimozkový hrbol – thalamus a okolní útvary
|
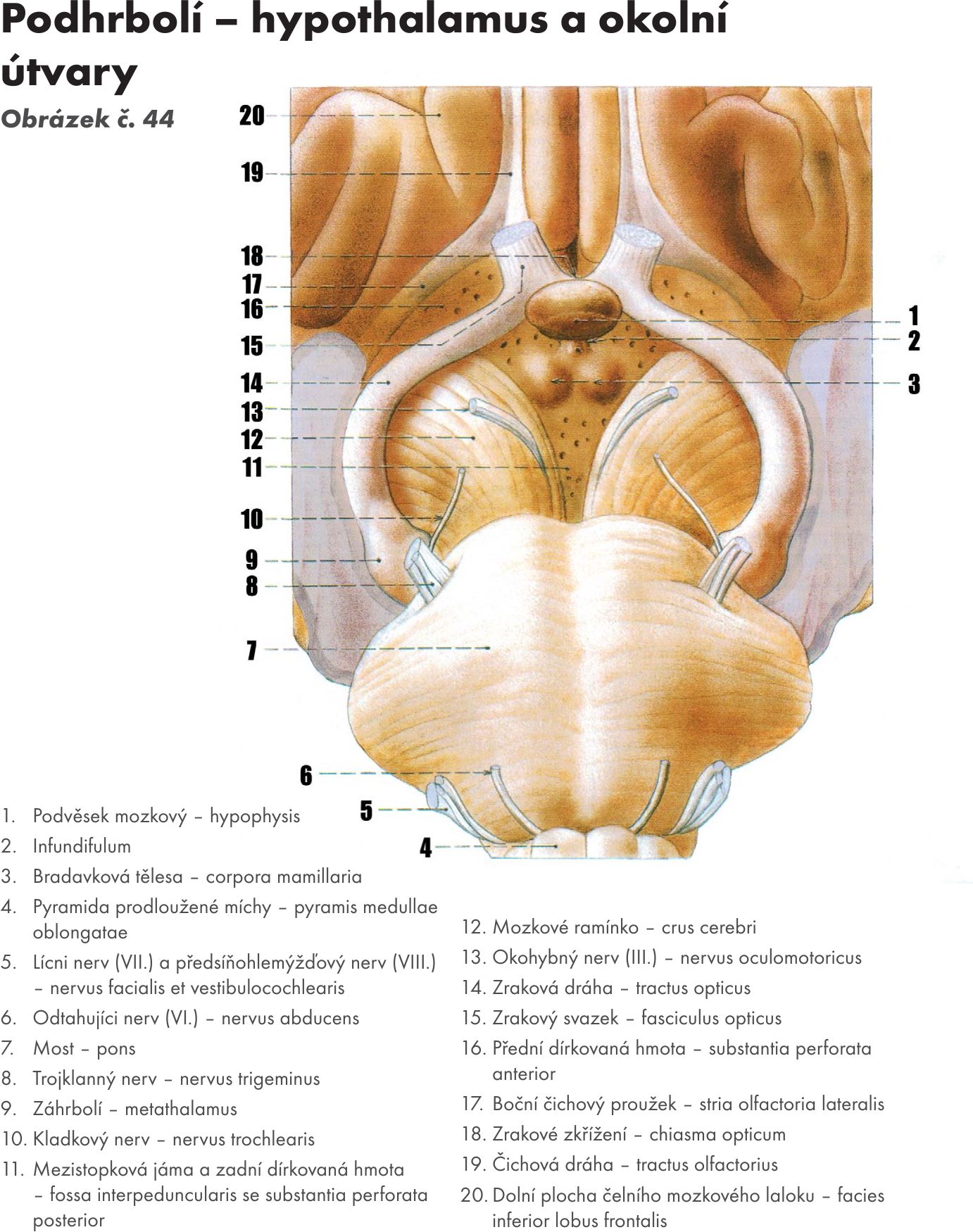
Obr. 44 Podhrbolí – hypothalamus a okolní útvary
|
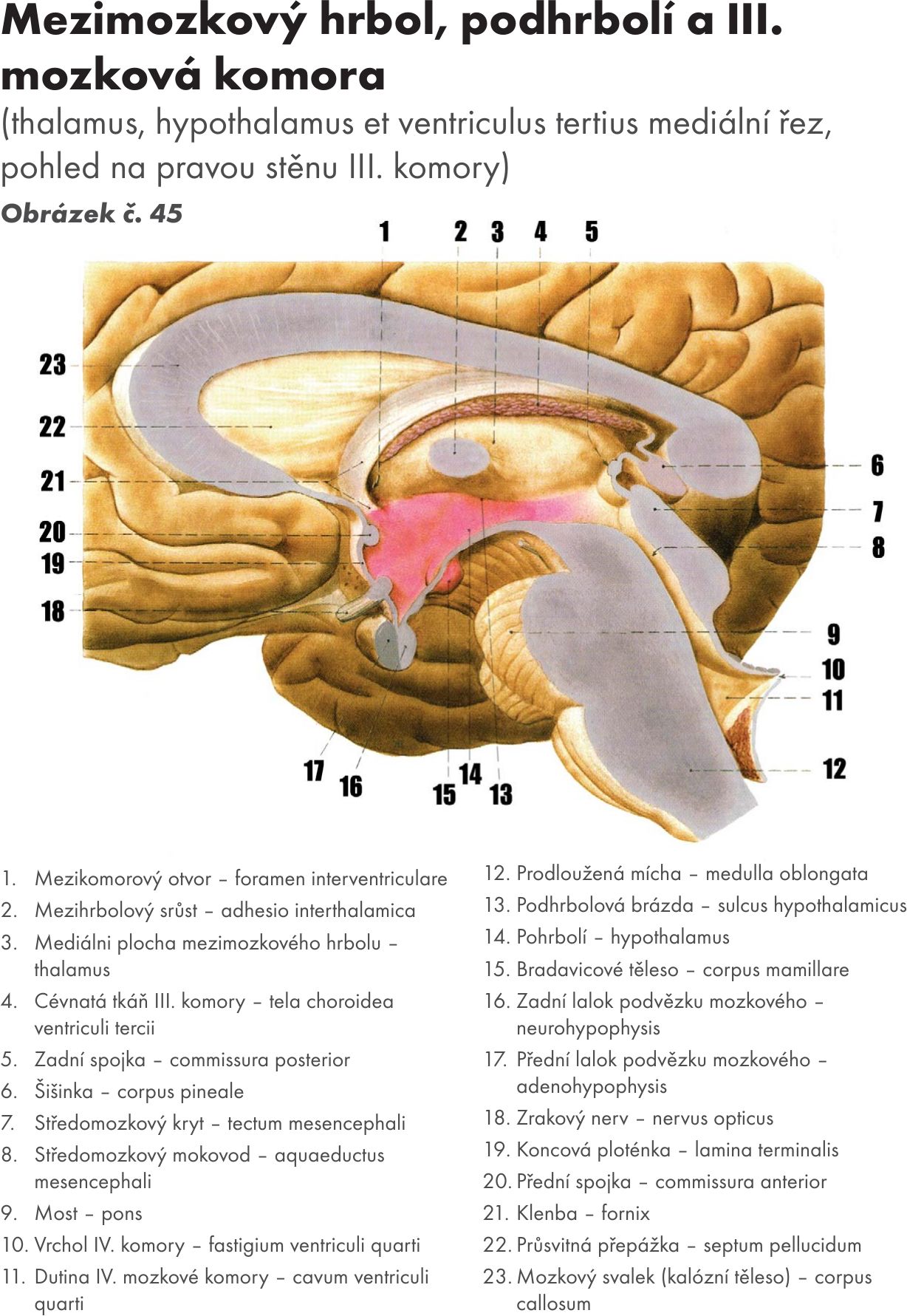
Obr. 45 Mezimozkový hrbol, podhrbolí a III. mozková komora
|
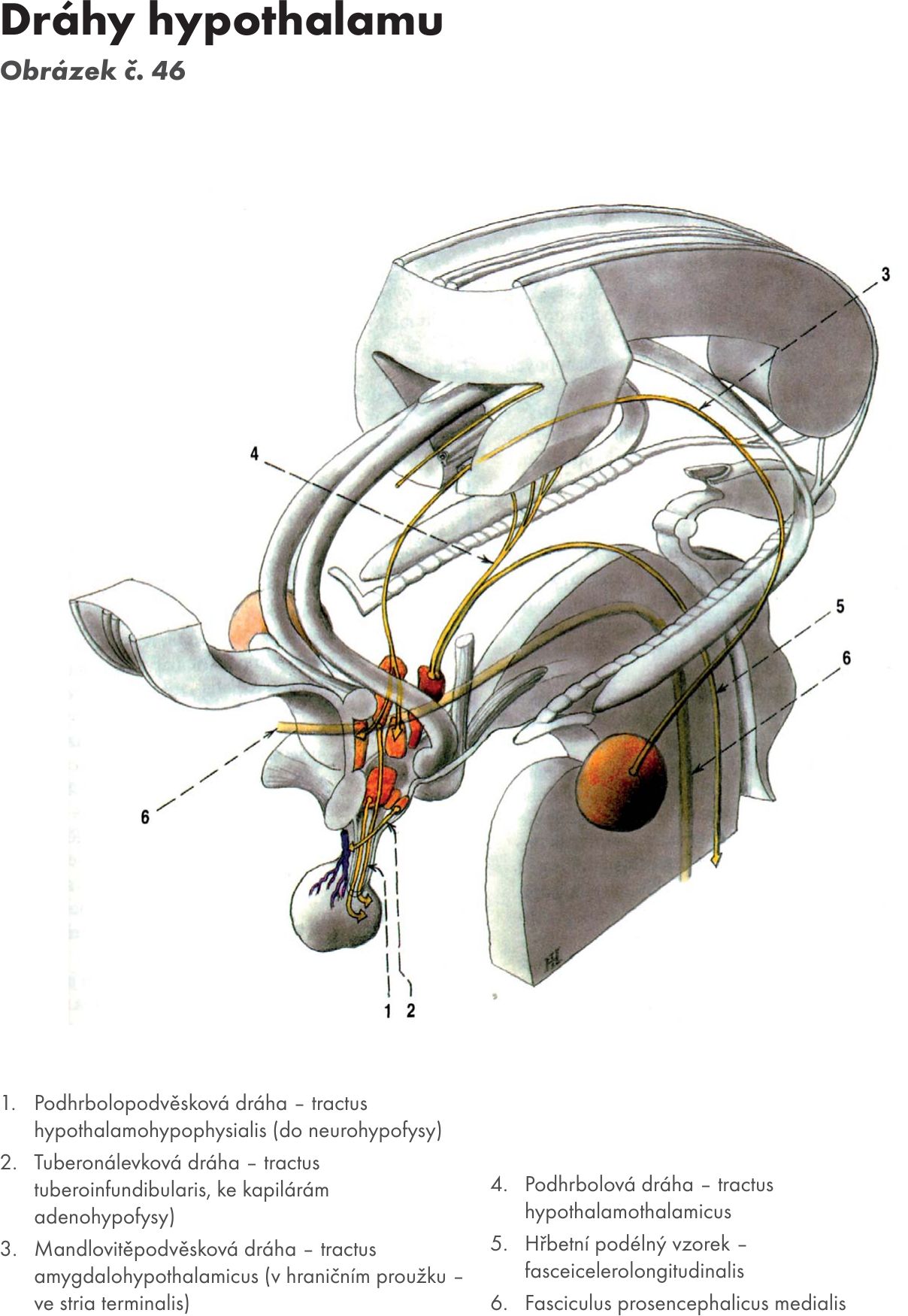
Obr. 46 Dráhy hypothalamu
|
|||
Koncový mozek – telencephalon
-
Základní členění
Je nejrostrálnější mozkový oddíl, který se skládá ze dvou hemisfér, které jsou od sebe odděleny sagitálně orientovanou štěrbinou – fissura longitudinalis cerebri, do které zasahuje srp tvrdé pleny mozkové – falx cerebri. Mozková kůra je řídící soustavou nervového systému a vykonává nejsložitější nervové funkce, které jsou spojeny s vědomím, inteligencí, instinkty atd. Díky ní můžeme komunikovat, pamatovat si a rozumět. Mozek je taktéž zahrnut do senzitivního,senzorického i motorického systému připojením na hlavové nervy a periferní nervy, cestou míchy, a kontroluje chtěné (volní) pohyby.Průněrmá hmotnost mozku je asi 1500 g.
Během embryonálního vývoje se v rostrální části nervové trubice již u 4 týdenního zárodku vytváří 3 primární mozkové váčky :
- přední mozek – prosencephalon
- střední mozek –mesencephalon
- zadní mozek – rhombencephalon.
Z kaudální části nervové trubice se vytvoří mícha, jejímž středem probíhá centrální kanál.V pátém týdnu se ze tří primárních váčků stává pět sekundární váčků.
- Přední mozek se rozděluje na:
- koncový mozek- telencephalon, ke kterému patří mozkové hemisféry a basální ganglia. Ohraničují postranní komoru..
- mezimozek- diencephalon, ze kterého vznikají tři hlavní struktury
- hrbol- thalamus
- podhrbolí – hypothalamus
- nadhrbolí- epithalamus – ohraničují střední komoru
- záhrbolí – metathalamus
- Střední mozek se nedělí, zůstává mesencephalon, kterým vede mozkový mokovod (Sylviův).
- Zadní mozek se rozděluje na:
- zadní mozek – metencephalon, ke kterému patří
- most – pons Varoli
- mozeček – cerebellum
- myelencephalon, ze kterého vzniká prodloužená míchy.
- zadní mozek – metencephalon, ke kterému patří
Tyto útvary ohraničují čtvrtou komoru.
V průběhu plodového období mozek rychle roste a nastávají proporcionální změny. Do třetího měsíce nitroděložního života je povrch hemisfér hladký – lisencephalie. U nižsích savců hladký povrch hemisfér přetrvává. Jak hemisféry rostou ohýbájí se do tvaru podkovy a překrývají střední mozek i mezimozek. Kolem 6 měsíce se povrch hemisféry skládá do záhybů – mozkové závity – gyri cerebri a hemisféry nabývají vzhled vlasškého ořechu – gyrencephalie. Toto zprohýbání umožňuje umístění většího počtu nervových buněk do menšího prostoru. V 8 prenatálním měsíci je většina významných brázd vytvořena a u novorozence je vývoj gyrifikace v podstatě ukončen.
-
Zevní popis mozkových hemisfér Obrázek č. 47 Obrázek č. 48 Obrázek č. 49 Obrázek č. 50 Obrázek č. 51
Při pohledu na konvexní plochu mozku vidíme přední, čelní pól – polus frontalis a zadní týlní pól – polus occipitalis a pět hlavních laloků, které jsou pojmenovýny podle kostí uložených nad nimi.
- Čelní lalok – lobus frontalis – je největším lalokem mozku. Je oddělen od temenního laloku – lobus parietalis centrální brázdou – sulcus centralis, která probíhá v čelní rovniě před ní je uložen předcentrální závit – gyrus precentralis. Brázdami – sulci je čelní lalok dále dělen na horní, střední a dolní čelní závit- gyrus frontalis superior, medius et inferior.
- Temenní lalok – lobus parietalis je od čelního laloku oddělen centrální brázdou, od týlního laloku – lobus occipitalis, temeno-týlní rýhou (brázdou) – sulcus parietooccipitalis. Za centrální brázdou je uložen zacentrální závit – gyrus postcentralis. Závity je temenní lalok dále dělen na horní a dolní temenní lalůček – lobulus parietalis superior et inferior.
- Týlní lalok – lobus occipitalis. Od temenního laloku jej odděluje příčná týlní brázda- sulcus occipitalis transversus. Na konvexitě jsou nepravidelné závity- gyri occipitales. Na mediální ploše se popisují hluboká ostruhovitá brázda – sulcus calcarinus a před ní klínovitá skupina závitů, klín- fundus. Je sídlem korových analyzátorů zrakové oblasti.
- Spánkový lalok – lobus temporalis. Od čelního a temenního laloku je oddělen hlubokou boční brázdou – sulcu lateralis (Sylvii). Brázdami je spánkový lalok dělen na horní, střední a dolní spánkový závit – gyrus temporalis superior, medius et inferior. Na horní ploše horního spánkového závitu jsou 2–3 příčně uložené závity – gyri temporales transversi. První z nich se nazývá Heschlův závit a je sídlem korového centra sluchu.
- Ostrovní lalok – lobus insularis – insula. Je pátým lalokem mozkové hemisféry, který je hluboko zanořen v zevní brázdě a tvoří část jejího dna. Obsahuje několik závitů (gyri insulae) a je pokryt částmi spánkového, temenního i čelního laloku – tzv. víčky – čelotemenním a spánkovým – operculum frontoparietalis et temporalis.
-
Mozková kůra – cortex cerebri – šedá hmota
Je tvořena šedou hmotou, obsahuje těla nervových buněk, jejich dendrity, krátké nemyelinizované axony a neobsahuje nervové dráhy. Je tenká 2–4 mm, ale její plocha je trojnásobně zvětšena velkým množstvím závitů. Povrch kůry lidského mozku je asi 0,25 m2. Představuje přibližně 40 % celkové hmoty mozku. Všechny nervové buňky v mozkové kůře jsou interneurony, které jsou rozmístěny po celé CNS.
Ze studií struktury, zapojení a funkčních charakteristik se mozek člení na : starou kůru (allocortex), přechodnou kůru (mesocortex) a novou kůru (neuocortex).
Nová kůre – neocortex. Nejvíce je rozvinuta v mozku primátů a mozku lidském. Miliardy nervových buněk jsou sekupeny do šesti vrstev:
- vrstva (lamina zonati) obsahuje jen malé množství neuronů
- vrstva (lamina granularis externa) obsahuje hvězdicovité granulační a malé pyramidové neurony
- vrstva (lamina pyramidalis externa) obsahuje převážně menší pyramidové neurony. Neurony II. a III. vrstvy jsou hlavním zdrojem asociačních a komisurálních vláken.
- vrstva (lamina granularis interna) obsahuje hvězdicovité neurony. Na nich končí většina talamokortikálních vláken.
- vrstva (lamina pyramidalis interna) obsahuje převážně pyramidové neurony. Je hlavním zdrojem sestupných (descendentních) vláken, která končí na podkorových strukturách (basální ganglia, kmen mozkový, mícha).
- vrstva (lamina multiformis) obsahuje převážně vřetenovité buňky. Zde začíná většina kostikotalamických vláken.
Neocortex není homogenní struktura, mezi jeho oddíly existují významné rozdíly po celém povrchu mozkových hemisfér. Strukturní rozdíly byly podkladem pro cytoarchitektonické členění mozkové kůry. Nejúspěšnější je Brodmanova mapa, který v roce 1909 rozdělil kůru do 52 funkčně odlišných oblastí.
-
Korové analyzátory Obrázek č. 52 Obrázek č. 53
V mozkové kůře se nacházejí tři typy funkčních oblastí, ve kterých jsou uloženy korové analyzátory:
- Pohybové oblasti (motorické), které kontrolují chtěné pohyby a jsou ukloženy v zadní části čelního laloku. Patří k nim:
- Primární motorická (somatická) kůra. Odpovídá 4. Brodmanově oblasti. Nacházejí se zde hlavně velké pyramidové buňky. Jejich axony tvoří mohutnou pyramidovou dráhu (koromíšní – tractus corticospinalis, viz motorická dráha, str.) Axony buněk se kříží, takže levá primární motorická kůra kontroluje svaly na pravé straně těla a naopak. Dráha zajišťuje přesné a zručné volní pohyby těla, zvláště předloktí, prstů a tvářových svalů. Prostorové uspořádání pyramidových buněk, které kontrolují jednotlivé svaly znázorňuje Obr. Mapka pro motorickou kůru se nazývá motorický homunkulus (tzv. človíček).
- Premotorická kůra. Odpovídá 6. Brodmanově oblasti. Tato oblast kontroluje složitější pohyby, přijímá specializované senzitivní informace (zrakové, sluchové, senzitivní podněty těla z ostatních oblastí kůry.
- Frontální zrakové pole. Odpovídá 8. Brodmanově oblasti. Kontroluje chtěné rychlé pohyby očí – sledování pohyblivého cíle.
- Brocova oblast. Překrývá 44. a 45. Brodmanovu oblast uloženou v dolním závitu čelního laloku – gyrus frontalis interiér a v čelním víku – operculum frontalis. Řídí tvorbu řeči, kontroluje pohyby nutné pro mluvení a emotivní zabarvení mluveného slova, podílí se na uchování pracovní paměti slov a řeči (tzv. krátkodobá paměť). U praváků je oblast v levé hemisféře.
- Senzitivní oblasti – podílejí se na smyslovém vnímání (zrak, sluch, rovnováha, chuť….) a jsou uloženy v temenním, spánkovém a týlním laloku.
- Primární somatosenzitivní kůra. Odpovídá 1.–3. Brodmanově oblasti. Je zodpovědná za vnímání obecných tělesných vjemů: bolest, tlak, vibrace a propriocepce (= vnímání jednotlivých částí těla) v kůži, tělesné stěně a končetinách. Tyto senzitivní informace jsou zachycovány senzitivními receptory a přenášeny míchou a mozkovým kmenem do somatosenzitivní kůry (str …). Schopnost lokalizovat přesně stimuly se nazývá prostorová diskriminace (určení rozdílů). Pravá mozková kůra přijímá senzitivní informace z levé části těla a naopak. Mapka pro senzitivní kůru se nazývá –senzitivní homunkulus (tzv. človíček). Největší počet senzitivních receptorů je obsažen ve rtech a konečcích prstů. Poškození primární senzitivní kůry naruší schopnost cítění bolesti a teploty, také lokalizaci dotyku, tlaku a vibrací na kůži.
- Somatosenzitivní asociační oblast. Odpovídá 5. a 7. Brodmanově oblasti. Mají spoje s primární somatosensorickou kůrou a umožňuje rozlišení jednlotivých senzitivní podnětů (hmat, tlak…) na základě uchované paměti dřívějších senzitivních vjemů a zkušeností.
Pozn.: když sáhneme do kapsy, somatosenzitivní oblast nám umožňuje zjistit, jaký předmět cítíme (klíče, mince, pero…).
- Primární zraková oblast. Odpovídá 17. Brodmanově oblasti. Nachází se v zadní a vnitřní části týlního laloku v tzv. ostruhové brázdě – sulcus calcarinus. Přijímá zrakové informace, které vznikají na sítnici oka. Pokud je tato korová oblast zničena, člověk je funkčně slepý, i když oko zůstává nepoškozeno (není schopen uvědomovat si zrakové vjemy). Získané informace jsou zpracovávány na poměrně nízké úrovni (zpracovává informace z obou očí dohromady, vnímání orientace objektů..). Informace z pravé poloviny zrakového pole se dostávají do levé části zrakové kůry a naopak.
- Asociační zraková oblast. Odpovídá 18. a 19. Brodmanově oblasti spolupracuje s primární zrakovou oblastí a pokračuje ve zpracování zrakových informací určením barvy, tvaru a pohybu. Zrakové informace se převádějí dopředu přední a zadní dráhou, které se podílí na pojmenování „co“ a „kde“ se nachází.
- Primární sluchová oblast. Odpovídá 41. a 42. Brodmanově oblasti. Nachází se na horním okraji spánkového laloku (Heschlův závit). Po podráždění zvukových receptorů zvukovými vlnami jsou impulsy přeneseny do primární zvukové kůry, kde je rozpoznána hlasitost, rytmus a výška tónů (vysoké a nízké tóny).
- Asociační sluchová oblast. Odpovídá 22. Brodmanově oblasti a je uložena hned za primární sluchovou oblastí. Je odpovědná za rozeznání typu zvuků – hřmění, vřískot, hudba atd. a je zde uložena paměť zvuků. Leží zde funkční mozkové oblasti pro rozeznávání a porozumění jednotlivým slovům (Wernikeovy řečové oblasti). Jejich poškození má za následek ztrátu schopnosti rozumět mluvenému slovu – senzorická afázie.
- Chuťová kůra. Odpovídá 43. Brodmanově oblastí a je uložena na vrcholu boční brázdy – sulcu lateralis. Umožňuje vnímání chuťových vjemů.
- Vestibulární kůra. Je uložena v zadní části ostrovního laloku – insula a je odpovědná za vnimání rovnováhy, tzv. polohy hlavy v prostoru.
- Čichová kůra. Odpovídá 28. a 34. Brodmanově oblasti a je uložena na vnitřní straně mozku, spánkového laloku, v hruškovitém laloku – lobus piriformis v oblasti hákovitého laloku – uncus. Čichové impulsy z nosní dutiny (pars olfactoria) sem přivádí čichové nervy (I. hlavový nerv – čichová dráha). Čichová kůra je částí nosního mozku – rhinencephala a spojuje se s mozkovými oblastmi, které vnímají emoce – limbický systém, proto vůně často vyvolávají emoce. Část čelního laloku nad očnicí – orbitofrontální kůra umožňuje identifikaci, rozpoznání na specifické vůně, zápachy a jejich pojmenování.
- Asociační oblasti jsou to oblasti s vyšším řádem zpracování. Spojují různé informace a umožňují tak smysluplnou akci.
- Předčelní kůra – cortex prefrontalis. Je uložen před motorickými oblastmi a podílí se na poznávacích (kognitivních) funkcích. Zahrnuje všechny pochody vnímání, myšlení, vědomí, zapamatování si s rozpomínáním se na informace. Je nutná pro abstraktní myšlení, sebekontrolu, uvažování a úsudek, houževnatost, řešení složitých problémů, dlouhodobé plánování, přizpůsobivost, společenské chování, smysl pro humor, schopnost vcítění se a svědomí. Má spojení s limbickou částí předního mozku a tím také podíl na náladě a emocích. Podílí se na paměti prostorových vztahů, pracovní paměti pro objekty a obličeje. Poranění této oblasti způsobuje těžké deprese.
- Všeobecná analyzační oblast. Oblast pro všeobecné vyhodnocení. Nachází se v zadní a zevní části mozkové kůry a je ve vzájemném kontaktu se zrakovou, sluchovou a somatosenzitivní oblastí a všechny typy těchto senzitivních informací spojuje. Poškození této oblasti vyvolává agnoziíí (ztrátu schopnosti vnímat, rozeznávat předměty, osoby). Na spodině spánkového laloku se nachází vřetenovitý závit – gyrus fusiformis, který spojuje vizuální, paměťovou a rozlišovací schopnost pro obličej. Při poškození této oblasti vzniká porucha tzv. faceblind, porucha slepého obličeje, prosopagnosie. Takto postižení lidé rozeznávají druhé lidi podle účesů.
- Řečová oblast. Rozsáhlá oblast obklopující boční brázdu Sylvickou – sulcus lateralis Sylvii v levé mozkové hemisféře. Zahrnuje různé funkce spojené s řečí. Dosud je identifikováno 5 oblasti:
- Brocova oblast – tvorba řeči,
- Wernickeova oblasti – porozumění řeči
- boční prefrontální kůra těsně před a pod Brocovou oblastí – podrobná analýza mluveného slova
- většina oblasti zevní a dolní části spánkového laloku, koordinace sluchových a zrakových stránek řeči, jako je pojmenovávání viděných objektů a slyšených slov
- části inzuly – spuštění slovní artikulace a rozpoznání rytmu a zvuku mluveného slova.
Dále je to paměťová oblast pro slova (z toho vyplývají i typy poškození řeči – fatické poruchy – motorické, senzitivní, mnestické).
Tyto oblasti v pravé hemisféře se nepodílejí na tvorbě řeči, spolupracují při výkladu slov a kontrole emočního podbarvení řeči.
- Insula. Některé jejích oblasti se podílí na tvorbě řeči, na rovnováze a na orgánových funkcích, včetně vědomého vnímání vjemů z orgánů (plný močový měchýř, nevolnost žaludku, některé vůně). Má vliv i na chování srdečně cévního systému (chvilkové „zastavení“ srdce polekaného člověka).
Typy agnosie19
název
popis
Akinetopsie
Cerebrální akinetopsie je spojena s neschopností vnimat zrakem pohyb. Cerebrální akinetopsii způsobuje léze vně striatové kůry
Anosognosie
Je neschopnost mít zpětnou vazbu o svém vlastním stavu a může být zaměněna s nedostatkem náhledu, ale příčinou je problem ve zpětnovazebném mechanismu mozku.
Aperceptivní visualní (zraková) agnosie
Problém s rozlišením vizuální ostrosti s problém s rozpoznáním, kopírováním nebo rozlišením mezi různými visualními stimuly.
Asociativní visualní (zraková) agnosie
Pacient může pospat vizuální scény a zařazovat objekty, ale selhává při jajich rozpoznání. Např. ví co je vidlička, ví že se s ní jí, ale nemůže ji rozlišit od lžíce. Patient s asociativní agnosií je ještě schopen reprodukce obrazu cestou kopírování.
Astereognosis
Somatosensorická agnosie je spojena s dotykem. Pacient má problém rozlišit objekty hmatem na základě jejich textury, velikosti a váhy. Může je pospat verbálně nebo je rozpoznat z obrázku či kresby. Vychází z poškození v somatosensorickém kortexu.
Auditorní (sluchová) agnosie
Auditorní (sluchová) agnosie byla popsána v roce 1877. Se sluchovou agnosií je obtížné porozumění zvukům z prostředí a neverbalním sluchovým impulsům včetně obtížného rozlišení hlasového a nehlasového zvuku, přestože je sluch normální. Jsou dva typy: semantická asociativní a discriminativní agnosie. Semantická asociativní agnosie je spojena s postižením levé hemisféry, zatímco discriminativní agnosia je spojena s lezí pravé hemisféry.
Auditorní (sluchová) verbální agnosie
Čistá slovní hluchota (Pure Word Deafness – PWD) představuje formu hluchoty s intaktním sluchovým ústrojím, ale signifikantně je obtížné rozeznat mluvená slova v jejich semantickém smyslu.
Autotopagnosie
Je to neschopnost orientace částí těla. Častou příčinou je poškození v parietalní oblasti zadního hrbolového paprsčitého vláknění – radiationes thalamicae posteriores.
Cerebrální achromatopsie
Barevná agnosie je postižení rozlišení kategorizace barev a vlastních barev.2 oblasti mozku se specializují na barevné vnímání a rozlišení (area V4 a V8). Jestliže je jednostranná léze v oblasti V4, pak je ztráta barevné percepce známá jako hemiachromatopsia.
Kortikální hluchota
Nerozlišujeme a nevnímáme žádné sluchové informace při nepoškozeném sluchovém ústrojí.
Environmentální agnosie
Je neschopnost lokalizace speciálně místností či budov, i když je důvěrně známe. Neschopnost stanovit trasu, jak se dostat či dojet do určité oblasti. Je obtížné na základě individuálních zkušeností se naučit cestu.. Tato forma agnosie bývá často spojena s poškozením oboustranným nebo pravostranným zadních oblasti hemisfér. Je často spojena s prosopagnosií a Parkinsonskou chorobou.
Prstová agnosie
Neschopnost rozlišit prsty na ruce. Je způsobena lezí v oblasti pareitálního loaku dominantní hemsféry a je součástí tzv. Gerstmannova syndromu má 4 primání symptomy:
- Dysgrafie/agrafie: deficit schopnosti psát
- Dyskalkulie/akalkulie: obtíže s učením nebo porozuměním matematice
- Prstová agnosie: neschopnost rozlišit prsty ruky
- Levo-pravá desorientace
Tvarová agnosie
Je neschopnost rozlišit detail, nikoli celý objekt.
Integrativní agnosie
Je formou asociativní agnosie nebo aperceptivní agnosie. U integrativní agnosie je pacient mezi těmito typy. Je to neschopnost rozlišit součásti něčeho a navíc neschopnost integrovat tyto prvky dohromady do srovnatelného vnímavostního celku.
Agnosie bolesti
Analgesia, je porucha vnímání a rozlišení bolesti; je to podklad sebepoškozování.
Phonagnosie
Neschopnost rozpoznat známé hlasy a navíc skrze sluch, jako posluchač nemůže rozumět užitým slovům
Prosopagnosie
Faceblindness a faciální agnosie: Neschopnost rozlišit a rozpoznat známé obličeje, rovněž včetně svého. To je mnohdy špatně presentováno jako neschopnost pamatovat si jména.
Čistá alexie
Neschopnost rozeznat text. Pacient s čistou alexií má často poškozené corpus callosum, nebo levou visuální asociační oblast. Čistá alexie je neschopnost číst tištěný materiál, ale někdy individuálně je ještě postižená schopnost psát.. Osoby s čistou alexií čtou slova po písmenech. Osoby s čistou alexií mají tzv. frekvenční efect. Jsou schopny číst slova s vysokou frekvencí lépe a rychleji než slova s nízkou frekvencí v textu.
Semantická agnosie
Je formou agnosie, kde prakticky je „slepý objekt“, využívají k rozpoznání nevizuální sensorické systémy, takže rozeznávají objekt např. pomocí pohmatu, poklepu, čichání, třesení nebo kmitání předmětu, mohou tak spustit realizaci jejich sémantického vjemu.
Sociálně emotionální agnosie
Je řazena mezi expresivní agnosie, je to forma agnosie, kdy osoba není schopna přijímat neverbální komunikační prvky – faciální exprese, řec těla a intonace řeči, ztvárnění, prorucha neumožňuje vnímat emoce a limituje aspekty sociálních interakcí.
Taktilní agnosie
Zhoršená schopnost rozpoznat a identifikovat objekty pouze hmatem
Časová agnosie
Je ztráta srovnávání sledu a průběhu událostí
Topografická desorientace
Topografická agnosia nebo topographagnosia, je forma zrakové agnosie, kdy osoba se nemůže spolehnout na zrakový podnět jako svého průvodce přímo v důsledku neschopnost rezpoznat objekty. Přesto mohou mít ještě excelentní schopnost popsat zrakové vrstvy ve shodném místě. Patient s topografickou agnosií má schopnost číst mapu, ale ztratí se v neznámém prostředí.
Vizuospatiální (zrakovoprostorová) dysgnosie
Je ztráta vnímání „kde je“ ve vztahu k sobě a prostředí a ve vztahu objektů k jiným objektům. To může zahrnovat i konstrukcionální apraxii, topografickou desorientaci, optickou ataxii, okulomotorickou apraxii, oblékací apraxii a pravo-levou záměnu
Vizuální agnosie
Je spojena s lezí levého okcipitálního laloku a spánkového laloku. Mnoho typů visuální agnosie obsahuje neschopnost rozpoznat objekt.
19 Agnosia. In Wikipedia, The Free Encyclopedia. Dostupné z WWW: http://en.wikipedia.org/w/index.php?title=Agnosia&oldid=608314880
Apraxie (z řeckého praxis = činnost) je charakterizována ztrátou schopnosti vykonat nebo naučit se cílené pohyby, přesto že chce a jsou fyzikální schonposti pohyby vykonat.. Je to porucha plánování pohybu, jež může být získaná či vrozená, ale příčinou není nekoordince, ztrata vnímání, či neschopnosti pochopit jednoduché příkazy. Příčinou jsou poškození speicfických oblasti mozku. Pozor na rozdíl mezi apraxií a ataxií, jako poruchou koordinace pohybu. Afázie je neschopnst produkce a pochopení jazyka; abulie, chyba v chuti vykonat pohyb; allochiria je stav, kdy osoba přijímá stimuli na jedné straně těla, které se ale stanou na jiné straně
Typy apraxie:
- Ideomotorická apraxie: jedná se o deficit schopnosti plánování a plné pohybové akce, která se spoléhá na sémantickou paměť. To může objasnit jak udělat pohyb, ale nemůže si jej představit nebo pohyb předvést, např. “ předstírat čistění zubů” či “ svraštit obličej při ochutnání citronové šťávy”, V případě automatického pohybu je schonpost intaktní. To je známo jako automaticko-volní dissociace. Např. nemůže zvednout telefon, když si řekne, ale může jej zvednout když chce zavolat.
- Ideacionální konceptuální apraxie: není schopnost vnímání úlohy a je narušená schopnost úplného mnohostupňovitého pohybu. Obsahuje schopnost selekce a vykonání příslušného pohybového programu. Např. lze provést plnou akci v nekorektní posloupnosti, jako natřít máslo na toast před vložením do opékače, nebo nazout si boty, před nazutím ponožek. Je to ztráty schopnosti vůle vytvořit naučenou úlohu, při využití nezbytných objektů a nástrojů. Např. vezme šroubovák a s ním zkusí psát jako s perem, nebo si zkusí učesat vlasy s kartáčkem na zuby.
- Bukofaciální nebo orofaciální apraxie: neverbální orální nebo bukofaciální ideomotorická apraxie, jejímž výsledkem je obtížné vykonání pohybů obličeje na požádání. Např. neschopnost olíznutí jednoho rtu nebo zapískat.
- Konstrukcionální apraxie: neschopnost nakreslit nebo zkonstruovat jednoduché konfigurace, např. křížící se pentagony.
- Apraxie chůze – porucha chůze
- Apraxie pohybu končetiny: obtíže s precisním pohybem horní či dolní končetiny.
- Okulomotorická apraxie: obtíže s pohybem očí, zejména se sakadovitým pohybem při přímém pohledu na cíl. Je to jedna ze 3 základních částí tzv. Balintova syndromu : neschopnost vnímání zrakového pole jako celku (simultanagnosia), obtížnost fixace očí (okulární apraxie), a neschopnost pohybovat rukou ke specifickému objektu díky zrakové kontrole (optická ataxie).
- Apraxie řeči: obtíže s plánováním a koordinací pohybu nezbytných k mluvení.20
20 Apraxia. In Wikipedia, The Free Encyclopedia. Dostupné z WWW: http://en.wikipedia.org/w/index.php?title=Apraxia&oldid=609106311
Afasia (pron.: /əˈfeɪʒə/ buď /əˈfeɪziə/ nebo /eɪˈfeɪziə/, z straořeckého ἀφασία (ἄφατος, ἀ- + φημί), "ztráta řeči” – němota) je porucha chápání a formulování jazyka kde příčinou je dysfunkce ve specifických oblastech mozku.
Tato skupina jazykových poruch obsahuje obtíže s pamatováním slov až po kompletní neschopnost mluvit, číst, psát. Jde vždy o ovlivnění visuálního jazyka, tak i znakového jazyka. Afasie je vždy spojena s poškozením mozku, velmi často mozkovou příhodou. Mozkové poškození, které způsobuje afazii může být vždy příčinou mnoha chorob mozku (nádory, padoucnice, Alzheimerova choroba apod.).
Akutní afazie se vyvíjí rychle jako výsledek poranění hlavy nebo mozková příhoda. Progresivní afázie se vyvíjí pomalu např. růstem nádoru mozku, infekcí, demencí. Oblast a rozsah postižení mozku nebo atrofie determinuje typ afázie a její příznaky..(afázie expresivní, receptivní, conduktivní, anomická, globální, primární progresivní a další.)
Klasifikace:
Klasifikace jednotlivých typů afázie je velmi obtížná. Problém je v neurální organizaci jazykové funkce v oblasti mozkové kory. Jen asi 60 % poruch lze na základě klasifikačního schématu zařadit,např. plynulá – neplynulá – čistá afázie. Některé typy jsou více selektivní – anomická afázie – deficit názvů, pojmenování, někdy jen pro některou skupinu objektů.
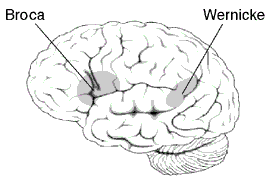
Lokalizační model primárních řečových center
Dle lokalizačního modelu vznikají 2 základní skupiny, dle časných studií Paula Broca a Carla Wernicke.
- expresivní afásie (Brocova afázie) – poškození centra v oblasti levého čelního laloku v oblasti motorického asociačního kortexu Brocovi oblasti. Osoby s tímto typem afázie mají často pravostrannou parésu či plegii ruky a nohy.
- sensorická afázie (Wernickeova afázie) – poškození spánkového laloku a vzniká fluetní – plynulá afázie, někdy označení receptivní afázie. Osoby s tímto postižením nemají žádnou tělesnou paresu, protože poškození je mimo oblasti motorické. V tomto typu afázie se dle lokalizace rozlišuje 5 typů:
- sluchová verbální agnosie(Pure Word Deafness = PWD)
- kondukční afázie
- řečová apraxie (nyní patří k samostatným poruchám)
- transkortikální motorická afázie (adynamická afázie či extrasylviánská motorická afázie)
- transkortikální senzorická afázie
- Anomická afáze, čili anomie nebo dysnomie, je typ afázie, kdy je potíž s pojmenováním objektů, někdy pouze selekce pro některé skupiny objektů.
- Globální afázie je výsledkem postižení rozsáklé oblasti kolem laterální rýhy – Sylvické brázdy – je problém s porozuměním mluvenému i psanému jazyku, a mají obtíže s mluvením. Jedná se o velmi těžký typ afázie.
- Izolovaná afázie neboli smíšená transkortikální afázie je typ poruchy jazykové dovednosti, je to neschonpost porozumět, co bylo řečen nebo potíž s tvorbou smyslné mluvy bez dojemné schopnosti recitovat, co bylo řešeno a získání nových slov k presentaci. Tento typ afázie vzniká postižením mozku v izolovaných oblastech řečové mozkové kůry – oblast levého spánkového a temenního laloku v okolí Wernikeova centra. Dochází k vnímání slov, ale ne jejich pochopení, nelze vytvořit vlastní smysluplnou řeč.
Progresivní afázie
Primární progresivní afázie (PPA) je spojena s progresivnimi onemocněními a demencí (frontotemporální demence, Pickův komplex motorického neurálního onemocnění, progresivní supranukleární paresa, Alzheimerova choroba). Je charakterizována stupňující se ztrátou schopnosti pojemnovávat objekty, je problém porozumět řeči, může být i potíž s nalezením správných slov a tvorbou vět. Dělí se na 3 typy : progresivní non fluetní afázie (PNFA), sémantická demence (SD), logopedická progresivní afázie(LPA).
Progresivní žargonová afázie je plynulá (fluentní) nebo receptivní afázie. Řeč je nesrozumitelná, ale připadá, že chápe její smysl. Řeč je plynulá a nenucená s neporušenou syntaxí, gramatikou, ale je problém se selekcí podstatných jmen. Jsou tvořeny neologismy, perseverace s pokusem nahradit slova, ale nelze nalést zvuk. Dochází k subtituci stejně znějících slov, sémantickým či fonetickým náhradám.
Fluentní – plynulá, nefluetní – neplynulá a “čistá” afázie
Různé typy afázie se dělí do tříkategorií: pylnulá, neplynulá, čistá.
- Receptivní afáze, se vždy označuje jako plynulá, fluentní, jsou postiženy vstup a příjem jazyka, jsou potíže se sluchovým verbální porozuměním, příjmem slov, frází, vět, které říká někdo jiný. Řeč je plynulá, ale je obtížné spojování výstupu jazyka – jako parafázie. Přiklady fluetní afázie jsou receptivní afázie, transkortikální senzorická afázie, konduktivní afázie, anomická afázie.
- Expresivní afázie, se vždy označuje jako neplynulá – nonfluetní afázie, jsou potíže s artikulací, ale v mnoha případech je relativně velmi dobré slovně verbální vnímání. Příklady neplynulé afázie jsou: expresivní afázie, transkortikální motorická afázie, globální afázie.
- “Čístá” afázie je selektivní porucha ve čtení, psaní a rozpoznání slov. Tato porucha může být zcela selektivní, např. osoba může číst, ale ne psát, nebo je schopna psát,ale ne číst. Příklady čisté afázie jsou: čistá alexie, agrefie, sluchová verbální agnosie.
Afázie se ještě dělí na primární a sekundární kognitivní proces:
Primární afázie je problém kognitivních jazykových procesních mechanismů: transkortikální senzorická afázie, sémantická demence, řečová apraxie, progresivní nonfluetní afázie, expresivní afázie.
Sekundární afázie je výsledkem jiných problémů, jako paměti, poruchy pozornosti, vnimavostní problémy: transkortikální motorická afázie, dynamická afázie, anomická afázie, receptivní afázie, progresivní žargonová afázie, kondukční afázie a dysartrie.
Jsou vytvářeny kongnitivní neuropsychologické modely snažící se objasnit jazykové procesy.
Příznaky a symptomy
Dysartrie a apraxie jdou mnohdy u některých postižení dohromady..
- neschopnost rozumění jazyku
- neschopnost výslovnosti, ale pro svalové ochrnutí
- neschopnost spontánní řeči
- neschonpost tvořit slova
- neschopnost pojmenovávt objekty
- špatná výslovnost
- přemrštěná tvorba a používání vlastních neologismů
- neschopnost opakovat fráze
- trvalé opakovaní frází (echolálie)
- parapfázie (nahrazování písmen, slabik či slov)
- agramatismus (neschopnost mluvit v gramaticky korektní formě)
- dysprosodie (postižení modulace, přízvuku a rytmu)
- neúplné věty
- neschopnost číst
- neschopnost psát
- limitovaná verbální tvorba
- obtíže s pojmenováním
- řečová porucha
Sumarizace různých typů akutní afázie:
Typ afázie
reprodukce
pojmenování
Sluchové porozumnění
Plynulost
Receptivní afázie
lehké -střední
lehké – těžké
vadné
plynulé, parafazické
Transkortikální senzorická afázie
dobré
střední-těžké
špatné
plynulé
Konduktivní afázie
špatné
špatné
relativně dobré
plynulé
Anomická afázie
lehké
střední-těžké
lehké
plynulé
Expresivní afázie
střední -těžké
střední-těžké
lehká obtížnost
neplynulé, pomalé, namáhavé
Transkortikální motorická afázie
dobré
lehké-těžké
lehké
neplynulé
Globální afázie
špatné
špatné
špatné
neplynulé
Smíšená transkortikální afázie
střední
špatné
špatné
neplynulé
- Osoby s receptivní afázií mohou mluvit v dlouhých větách,které nemají smysl, a navíc s nadbytečnými slovy, a neustále vytváří nová slova – neologismy. Mají špatné sluchové vnímání a vnímání textu, plynulý, ale absurdní, orální i písemný projev. Mají velké problémy s porozuměním řeči své i jiných a často si neuvědomují své chyby.
- Osoby s transkortikální senzorickou afázií mají podobný deficit jako s receptivní afázií, ale jsou schopny reprodukce bez chyby.
- Osoby s konduktivní afázií mají deficit ve spojení mezi vnímáním řeči a produkcí řeči. Jedná se o poškození obloukového svazku – fasciculus arcuatus, který převádí informace mezi oblasti Wernickeovou a Brockovou. Podobné příznaky jsou při poskožení inzult nebo sluchového kortexu. Sluchové vnímání je blízké normálu a slovní projev je plynulý s občasnými parafazickými chybami. Schopnost reprodukce je špatná.
- Osoby s anomickou afázií velmi obtížně pojmenovávájí. Mají potíže s označením slov, spojení gramatických typů (obtížné jmenování sloves a žádných podstatných jmen) nebo se sémantickými kategoriemi (minimálně obtížné pojmenování slov vztahujícím se k fotografii) či obecnějším pojmenováním. Mají sklon produkovat gramaticky zcela prázdnou řeč. Sluchové vnímání je nepoškozeno. Anomická afázie je afázie vznikající u tumorů v jazykové oblasti, dále u Alzheimerovi choroby.
- Osoby s expresivní afázií často hovoří krátce, významné, smysluplné fráze produkují s velkým vypětím. Expresivní afázie je charakterizována jako neplynulá. Často opomíjejí malá slova jako "je", "a" apod.. Jsou schopny v různé šíři rozumět řeči jiných. Uvědomují si často své potíže a jsou silně frustrováni.
- Osoby s transkortikální motorickou afázií mají podobný deficit jako s expresivní afázií, krom reprodukční schopnosti, která je normální. Sluchové vnímání je všeobecně vpořádku při jednoduché konverzaci, ale rychle klesá u komplexnější komunikace. Je spojena s pravostranou hemiparesou, kterou si pacient v oblasti horní končetiny a obličeje zcela neuvědomuje.
- Osoby s globální afázií mají velké komunikační potížesa jsou extrémne limitovány v jejich schopnost řeči nebo vnímání jazyka. Mohou zcela být nonverbální, nebo pouze použivají obličejové výrazy a gestikulace ke komunikaci. Je to spojeno s oblastí pravého obličeje a horní končetiny s pravostrannou hemiparesou, kterou si pacient neuvědomuje.
- Osoby se smíšenou transkortikální afázií mejí podoné problémy jako u globální afázie, ale schopnost reprodukce je zachována.
Subkortikální afázie – charakteristiky a příznaky jsou závislé na straně a velikosti podkorové léze. Zahrnuje v podkoží hrbol- thalamus, vnitřní pouzdro – capsula interna a basální ganglia.
Příčiny afázie : léze v oblasti čelního, spánkového a temenního laloku Brockovy a Wernickeovy oblasti a neurálních drah mezi nimi (cévní mozkvé příhoda, úraz mozku, jiná poranění mozku, mozkové nádory, progresivní neurologická onemocnění – Alzheimerova choroba, Parkinsonova choroba, chronické neurologické onemocnění – epilepsie, migrény, zánětlivá onemocnění mozku – virová, jako vedlejší efetk některých léků a toxinů).21
21 Aphasia. In Wikipedia, The Free Encyclopedia. Dostupné z WWW: http://en.wikipedia.org/w/index.php?title=Aphasia&oldid=618569582
Alexia (alexia z řečkého ἀ, vyjadřující negaci a λέξις = "slovo"), či získaná dyslexie, u poškození mozku dochází k problémum se schopnosti číst. Také se nazývá slovní slepota, textová slepota nebo vizuální afázie.
Klasifkace:
Jsou 2 skupiny:
- První,hlavní skupina je tzv. centrální dyslexie, kam patří: povrchová dyslexie, sémantická dyslexie, fonologická dyslexie a hluboká dyslexie.
- Druhá, tzv. periferní dyslexie, čítá: neglect dyslexii, pozornostní dyslexii, a čistou dyslexii. Je rovněž označena jako alexie bez agrafie.
Klasifikace závisí i na anatomickém postižení a prezentují ji: agrafie (deficit psaní) a afázie deficit orálního jazyka) Alexie a agrafie jsou často spojeny s afázií, ale existují i jednotky bez afázie. Pokud afázie není pak je alexie kategorizována : 1) alexie bez agraphie (čistá alexie nebo slovní slepota) – poškození levého týlního laloku, 2) alexia s agrafií – spojeno s poškozením levého úhlového závitu – gyrus angularis
Alexie – typické je poškození dominantní hemisféry mozku, převážně levé v oblasti temenního a týlního laloku, které odpovídají za procesy aspektů sluchových, fonologických a vizuálních. Oblast spojení mezi týlním s spánkovým lalokem (nazývající se jako týlně spánkové spojení – junctio occipito-temporale) koordinuje informace jdoucí z vizuálních a sluchových procesorů kory a přidělují význam těmto stimulům.Alexie může být i po postižení dolní části čelního laloku. Postižení různých oblastí kory má za následek rozdílná schémata v tíži postižení. I mozková příhoda může být příčinou alexie.
Alexie bez agrafie je diskonekční syndrom, který se vyvíjí přerušením komunikační cesty mezi pravou a levou hemisférou. Pravý vizuální kortex přijímá informaci z levé strany a levého vizuálního pole a tato informace je vedena cestou zadní spojky – commisura posterior a zduřením mozkového svalku – splenium corporis callosi do slovotvorného systému v levé hemisféře. Tato cesta je v tomto případě přerušena.
Presentace agrafie je výsledkem léze zadní části levého čelního laloku a levé horní části temenního laloku.
Alexie může být spojena s expresivní afázií, nebo receptivní afázií. Alexie ve spojení s agrafií, může být specifická ztráta schopnosti produkce psaného jazyka, přestože motorické manuální schopnosti jsou intaktní. Jindy může být postižení oblasti, která odpovídá za vstupní proces. Výsledkem je známá čistá alexie. Jedná se o stav, kdy schopnost tvořit psaný jazyk je nahrazena neschopností rozumět psanému textu.
Strefosymbolie (Samula Orton, 1925) je odpovědí na důsledky tvorby slovní slepoty a jak stav kompezovat. Strephosymbolií je stav obtížné tvorby konců viděných napsaných slov u neznámých jazyků, zejména v případě, že písmena jsou vzhledově podobná jiným např. b-d, m-n, p-q.

Asfedia :je skotopický senzitivní syndrom nebo vizuální námahový syndrom,je znám jako Irlenův syndrom. Jedná se o zrakovou percepční vadu spojenou primárně se čtením a psaním. Může mít široký vliv na paměť a koncentraci.
Synestezie (z řeckého syn- = spojení a aesthesis = cítění, vjem) je sdružení vjemů dvou nebo více tělesných smyslů. Jedná se o způsob vnímání, kdy původní vjem jednoho smyslu vyvolá vjem smyslu jiného, resp. je k původnímu přiřazen. Může jít i o sdružení různých vjemů, které by patřily stejnému smyslu, avšak vyvolaný vjem je pouze „cítěn“. V obou těchto případech může být první či vyvolávající z vjemů viděn, slyšen, apod. nikoli fakticky, ale jen mentálně, tedy osoba si jej představuje ve své mysli.
Základní charakteristiky synestezie
- vnímání nastává nedobrovolně a spontánně
- vnímání je individuální: každý synestetik má "vlastní" barvy či tvary
- vnímání jsou nepřevoditelná: číslo sedm vyvolá modrou barvu, ale modrá barva nevyvolá číslo 7.
- schopnost je trvalá a neměnná.
Synestezie má mnoho podob a neexistuje kombinační strop pro její možnosti. K nejznámějším patří tato spojení:
- grafický symbol (např. písmeno nebo číslo) → barva, jakkoli oba vjemy patří jednomu, tedy zrakovému smyslu, vyvolaný vjem barvy je pouze „cítěn“. I první z nich může být ovšem pouze „myšlen“ (jak výše popsáno), nikoli skutečně okem zaznamenán.
- hudba, zvuk → barva, spojení akustického a optického vjemu se nazývá synopsie. Vybaví-li se vnímateli při poslechu hudby nebo zvuku vizuální vjem, jedná se o fotisma, opačný případ se nazývá fonisma. Fotismata jako první popsal ruský skladatel Nikolaj Rimskij-Korsakov, který přiřazoval barvy jednotlivým tónům (např. tón C = bílá, D = žlutá barva, atd. – viz autorova stránka). Francouzský skladatel Olivier Messiaen, který měl barvy spojené s tóninami i s hudebním výrazem: Říkal s oblibou interpretům v orchestru např: „Hrajte to víc zeleně“. Podobným způsobem mohou působit i vjemy nehudebních zvuků. Existují také zajímavé analogie v pojmovém označování (např. vysoký – nízký tón).
- personifikace, v tomto případě mohou být čísla, dny, měsíce či písmena asociovány s určitými osobami, tedy personifikovány.
- slovo → grafický vjem, zde určité slovo vyvolá nejčastěji představu geometrického obrazce či jiného útvaru.
- slovo → chuťový vjem, individuálně specifická slova mohou být spojena s pocitem určité chuti v ústech. Tato forma synestezie je řídká.
- zrakový vjem → hmatový vjem, pocit dotyku nebo ohmatávání viděného objektu, nebo celého zorného pole najednou.
U někoho se synestézie objevuje po celý život, někdo pozná tuto zkušenost jen po přechodnou dobu, u někoho nefunguje vůbec. Synestézie se podle epidemiologických studií projevuje asi šestkrát častěji u žen, takže úvaha o vazbě na chromozom X má smysl. Současné genetické výzkumy to potvrzují, a dokonce se vyskytují úvahy, že pokud by se objevil gen pro synestézii u plodů mužského pohlaví, mohl by být spojen s dalšími poruchami, které vedou k jeho časnému odumření a potratu. Mužské plody se potracují častěji. Nejvíce stop míří k chromozomu 16, přičemž genotyp se nemusí vždy „prosadit“, jak ukazují studie na jednovaječných dvojčatech. Vyskytuje se hojně u dětí a ustupuje v procesu ontogeneze. Synestézie jsou hojnější u přírodních národů, vyskytují se častěji u umělců než u jiných skupin lidí a jsou příznačné také pro některé psychopatologické fenomény (např. u schizofreniků). Schopnost vnímání tohoto typu má také částečně genetický podklad. Synestézie je potvrzený jev, ale způsob tohoto vnímání je ryze individuální, proto teorie, určující přesné přiřazení kvality vjemů nelze považovat za zcela relevantní.11
11 Wikipedie: Otevřená encyklopedie: Synestezie [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Synestezie&oldid=10987242
Fotisma je druh synestezie, při kterém je reálný akustický vjem doprovázen vnitřním optickým vjemem. Opačný druh je fonisma, při kterém je reálný optický vjem doprovázen vnitřním akustickým vjemem. Všechny typy spojení akustických a optických kvalit se nazývají synopsie.
Fotismatajsou nejčastější formou synestezií. Objevují se asi u 12 % lidí (fonismata asi u 4 %). Častou formou fotismat je „barevné slyšení“ (chromatická synopsie) – při poslechu vystupují intenzivní pestré barvy. Podle G. Anschütze lze rozdělit synopsie na analytické (spojení jednotlivých tónů a barev, u některých osob vznikají celé systémy fotismat), syntetickéči komplexní (spojení hudebních celků s klidnými nebo dynamickými obrazy) a analyticko-syntetické(barvy jsou přiřazovány tóninám). Ve spojování tónů a barev se vyskytují u některých osob nezávislé shody (např. spojování tónu f' či zvuku trubky s červenou barvou), většinou se však synopsie u různých lidí (i podstatně) liší.12
12 Wikipedie: Otevřená encyklopedie: Fotisma [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Fotisma&oldid=7386973
Synestézie [synesthesia] je vnímání jednoho smyslu druhým (např. slyšení barev, barevné vidění zvuku …). Věda sice přesně nezná důvody tohoto jevu, ale aspon jej nepopírá (jako u jiných věcí). Uvádí se, že tuto schopnost zažil aspon jeden ze dvou tisíc lidí a má ji 5 % dětí předškolního věku. Podle jiného zdroje jsou to 2 lidé z milionu. (obr.)
Jako příklad barevného slyšení se často uvádí básník Arthur Rimbaud (1854–1891), konkrétně jeho básen Voyelles (samohlásky).Ale i básník Charles-Pierre Baudelaire, nebo třeba americký fyzik Richard Feynman se jednou přiznal, že viděl matematické symboly Besselovy funkce jako barevné schéma. Velmi častý výskyt syntestézie je i mezi lidmi vlastnící absolutní sluch a jednotlivé tóny vidí jako barvy např. Franz Liszt, Franz Schubert, ale i u Ludwiga Beethovena jsou určité náznaky této psychologické schopnosti.
Syntestézii studoval a popisoval neurolog Vilayanur S. Ramachandran, který ji přirovnal k obecné schopnosti metafóry, kdy vlastně také nahrazujeme určité věci a stavy do jiných rovin než, kterým původně naleží.
Německý psycholog Wolgang Köhler zase navrhl psychologický experiment, který sice v původním záměru dokazoval způsob vývoje jazyka a mluveného slova, ale pro základní představu jak myslí a uvažují synestetici nám postačí. Podívejte se na dva obrázky a zkuste si tipnout, který tvar se jmenuje Booba a který se nazývá Kiki.
Kolem 97 % všech lidí označí oranžový tvar jako Kiki, kvůli ostrým a hranatým tvarům. Zaoblený tvar je vybrán mimo jiné, protože i ústa při vyslovování mají oblý tvar
Ve funkci pravé a levé hemisféry existují některé rozdíly. Obě hemisféry kontrolují opačné části těla. Asi u 90–95 % lidi kontroluje levá hemisféry spíše jazykové schopnosti, matematicku, pracuje s malými detaily, které potom logisky vysvětluje. Pravá hemisféry se podílí více na zrakově prostorových schopnostech, emocích, schopnostech uměleckých a hudebních, na rozeznávání výrazů obličeje. Přes rozdíly hemisféry spolu komunikují pomocí spojovacích vláken – komisur.
- Pohybové oblasti (motorické), které kontrolují chtěné pohyby a jsou ukloženy v zadní části čelního laloku. Patří k nim:
-
Bílá hmota mozková
Leží pod šedou hmotou mozkovou a tvoří ji vlákna a dráhy:
- Asociační vlákna. Spojují sousední korové oblasti v jedné hemisféře, krátká asociační vlákna – fibrae arcuatae nebo vzdálenější místa kůry – dlouhá asociační vlákna, která vytváření svazky.
- Komisurální (spojovací) . Komisury – spojky probíhají mezi oběma hemisférami a propojují odpovídající oblasti šedé hmoty – homotropní vlákna, nebo spojují různé oblasti levé a pravé hemisféry – heterotropní vlákna. Umožňují spolupráci obou hemisfér a jejich koordinací. Tvoří je:
- Mozkový svalek (svorník, kalózní těleso) – corpus callosum. Je největší mozková spojka – komisura, která vzájemně spojuje kůru frontálních, temenních a týlních laloků hemisfér. Z temporálních laloků spojuje pouze sluchové oblasti. Popisujeme na něm 4 části: svalkové zduření, kmen, koleno a zobák (splenuim, truncus, genu et rostrum corporis callosi).
- Přední spojka – commisura anterior. Leží před III.komorou. Spojuje čichové oblasti obou hemisfér – pars anterior a temporální laloky, kromě sluchových oblastí (pars posterior).
- Klenba – commisura fornicis. Leží pod svorníkem a spojuje hippokampové útvary (hippocampus, lůžko – subiculum, zubatý závit – gyrus dentatus). Popisujeme na něm 3 části: klenbové raménko, tělo a sloupek (crus, corpus et columnae fornicis).
- Projekční vlákna. Sestupují z mozkové kůry do nižších oblasti CNS nebo naopak. Díky těmto vláknům se dostávají motorické impulsy do periferie a senzitivní informace z periferie do kůry.
Probíhají vertikálně a dělí se na :
- Krátká projekční vlákna. Spojují mozkovou kůru s bazálními ganglii a mozkovou kůru s thalamem. Dráhy jdou z kůry i do kůry.
- Dlouhá projekční vlákna. Vytváření dráhy, které probíhají z kůry a hluboko v bílé hmotě mozkové tvoří kompaktní svazek – vnitřní pouzdro – capsula interna, který se nachází mezi thalamam a ocasatým jádrem (nukleus caudatus), které leží od něho na straně mediální a jádrem čočkovitým (nuclesu lentiformis), které leží od něho na straně laterální Nad capsula interna se projekční vlákna vějiřovitě rozbíhají v paprsčitý věnec – corona radiata. V capsula interna mají jednotlivé dráhy charakteristickou polohu.
Vnitřní pouzdro má 3 části: přední ramínko, koleno a zadní ramínko (crus anterius, genu, crus posterius capsulae internae).
- Přední ramínko - crus anterius capsulae internae obsahuje přední hrbolová paprsčitá vláknění – radiationes thalamicae anteriores, čelomostní dráhu – tractus frontopontinus,
- Kolénko – genu capsulae internae obsahuje korojádrovou dráha – tractus corticonuclearis,
- Zadní raménko – crus posterius capsulae internae se dělí na 3 části :
- hrboločočkovitou část – pars thalamolenticularis (obsahuje vlákna koromíšní – fibrae corticospinales, koročervené – fibrae corticorubrales, korosiťované – fibrae corticoreticulares, hrbolotemenní – fibrae thalamoparietales, centrální hrbolová paprsčitá vláknění – radiationes thalamicae centrales),
- podčočkovitou část – pars sublenticularis (obsahuje zrakové paprsčité vláknění – radiatio optica, sluchové paprsčité vláknění – radiatio acustica, korokrytová vlákna – fibrae corticotectales, spánkovomostní vlákna- fibrae temporopontinae) a
- začočkovitou část – pars retrolenticularis (obsahuje zadní hrbolová paprsčitá vláknění – radioationes thalamicae posteriores, temeno-týlní mostní svazek – fasciculus parietooccipitopontinus).
-
Basální ganglia Obrázek č. 54 Obrázek č. 55
Hluboko uvnitř bílé hmoty mozkové jsou uloženy objemné shluky šedé hmoty, uspořádané do útvarů – basální ganglia.
Patří k ním:
- Čočkovité jádro – nucleus lentiformis. Má dvě části: laterální – skořápka (putamen) a mediální bledá koule- globus pallidus. Leží laterálně od vnitřního pouzdra- capsula interna. Tenká vrstva bílé hmoty.rozděluje bílou kouli – pallidum na vnitřní a zevní – pallium internum a pallidus externum. Při poškození pallida dochází k omezení až zástavě pohybů, zástavě řeči, ospalosti až kataleptickému stavu. Při poškození putamen dochází k bezděčným pohybům v klidu i při pohybu, mizí ve spánku (chorea).
- Ocasaté jádro – nucleus caudatus. Má tři části: hlavu, tělo a ocas (caput, corpus et cauda nuclei caudati), které se obtáčejí kolem hrbolu – thalamu. Hlava je uložena před hrbolem – thalamem a svou mediální plochou se vyklenuje do předního rohu postranní mozkové komory. Ocas obtáčí zadní část hrbolu – thalamu a zasahuje do spánkového laloku. Leží mediálně od vnitřního pouzdra – capsula interna. Poškození ncl. cudatus se projeví jako pomalé kroutivé pohyby distálních části končetin, grimasami v obličeji, řeč je nejasná (athetosa).
- Proužkovité těleso – corpus striatum – nazývají se tak dohromady ocasaté těleso - nucleus caudatus a laterální část čočkovitého jádra – skořápka (putamen). Obsahuje dva základní typy neuronů.
- Malé projekční neurony, které končí v bledém tělese – globus pallidus a v černé hmotě- substantia nigra (efferentní spoje)
- Inteneurony, které se větví uvnitř striata. Velké interneurony jsou cholinergní, malé jsou GABAergní.
Striatum dostává aferentní spoje z mozkové kůry, z hrbolu thalamu, a z mozkového kemen. Eferetní spoje vystupují ze striata a končí v bledém tělese – globus pallidus a v černé hmotě – substantia nigra. Při snížení přísunu dopaminu do striata a poškození černé hmoty- substantia nigra (degenerací apod.) se objevují příznaky parkinsonismu (hyperkineze). Mimovolní pohyby jsou rychlé, neúčelné a nepravidelné pohyby mimického svalstva, jazyka a svalů horní končetiny. Degenerace corpus striatum eventuálně mozkové kůry způsobuje také Huntigtonovu chorobu. Projevuje se zvýšenou stimulací motorických aktivit a tak se končetiny nekontrolovaně třesou. Přičinou je genetický defekt DNA.
- Závora – claustrum. Je proužek šedé hmoty uložený mezi skořápkou – putamen a ostrovním lalokem (insula). Od putamina ji odděluje vnější pouzdro – capsula externa a od inzulární kůry krajní pouzdro – capsula extrema. Aferetní spoje dostává ze všech nekortikálních oblastí, eferentní spoje míří zpět. Basální část přijímá spoje z čichové korové oblasti.
- Mandlové jádro – nucleus amygdalae. Leží v hlouby temporálního laloku, těsně před cornu anterius postranní mozkové komory, a před hippocampem. Dělí se na skupinu jader kortikomediální, vývojově starší, která obsahuje řadu peptidů a jádra basolaterální , vývojově mladší, která obsahují monoaminy (noradrenalis, dopamin, serotonin). Anatomicky se mandlové jádro (amygdala) přiřazuje k basálním gangliím, svým funkčním významem patří k limbickému systému. Aferetní spoje přicházejí do amygdaly z kortikální kůry frontální, temporální, inzulární,z čichové korové oblasti) z basálních ganglií (těmito je transportován acetylcholin), z thalamu, hypothalamu a retikulární formace.
Eferetní spoje vydávají vlákna do thalamu, hypothalamu, basálních ganglií a kortikální kůry, dlouhé spoje končí v substatia nigra, v retikulární formaci a jádrech IX. a X.
Poškození amygdaly má za následek ztrátu emočního prožívání.
Dle funkčního zapojení lze k basálním gangliím přiřadit i další struktury: černá hmota- substatina nigra, podhrbolové jádro (ncl. subthalamicus.) Poškození ncl. subthalamicus vyvolává mimovolní pohyby hlavně proximální části končetin, připomínající létací pohyby (balismus).
Basální ganglia pracují jako komplexní nervová struktura, která spolupracuje s mozkovou kůrou při kontrole pohybů, mejí tlumící vliv na korové i podkorové motorické funkce. Začínají, zastavují a regulují intenzitu volních pohybů, vybírají příslušné svaly nebo pohyby pro určitý úkol a brzdí protichůdně působící svaly. Mohou rovněž kontrolovat rytmus, opakující se úkoly a spoluúčastní se na tvorbě návyků. Základní okruh zpojení basální ganglií je :
premotorická korová oblast
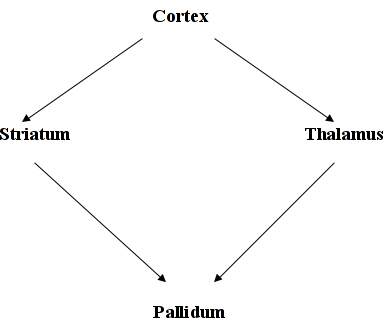
-
Limbický systém Obrázek č. 56
Limbická korová oblast je pruh kůry na mediální ploše hemisféry, obkružující mozkový svalek – corpus callosum. Počítáme k ní korové oblasti – kůra limbického laloku, jádra v telencephalu – amygdala, diencephalu, některá jádra thalamu a hypothalamu, mesencephalu, v pontu, aminergní jádra v telencephalu a mozkovém kmeni (jádra cholinergní, dopaminergní, noradrenergní a serotoninergní) a spoje limbického systému. Dále zahrnuje septální jádra, čelenkový závit – gyrus cinguli, podsvalkový závit – gyrus subcallosus, příhippocampový závit – gyrus parahippocampalis, část mandle –amygdaly a zanořené hippokampální formace (lůžko, podložka – subiculum, zubatý závit – gyrus dentatus, moršký koník – hippocampus). Zahrnuje rovněž malá jádra (basální Meynertovo jádro – cholinergní) a proužkovité jádro – corpus striatum (patří k basálním gangliím). V mezimozku jsou s limbickým systémem úzce spojena přední thalamická jádra a hypothalamus, cholinergní a aminergní jádra telencephala a kmene. Limbický systém také překrývá čichový mozek – rhinencephalon. Komisurální spoje limbického systému zajišťuje spojka klenbová – commissura fornicis a zadní části spojky přední – commissura anterior.
Všechny tyto struktury se podílí na vytváření oddílu předního mozku – oddíl prosencephalický a středního mozku – oddíl mesencephalický.
Oddíl prosencephalický má část korovu a podkorovou. V korové části rozlišujeme vnitřní a zevní okruh. Útvary těchto okruhů se účastní na vytvoření funkčně důležitého Papezova okruhu (Papez circuit). Obrázek č. 57
Základní schéma : fornix hippocampi → corpus mamillare → ncl. anterioris thalami → gyrus cinguli → gyrus parahippocapalis → hippocampulní formace.
Hlavní eferetní dráhy jdou do asociační kůry, do thalamu, hypothalamu a tegmenta.
Původní zapojení Papezova limbického okruhu v blokovém diagramu dle Domesicka (1972). Tučné linie – původní představa Papeze z r. 1937 (Stephan, 1975)
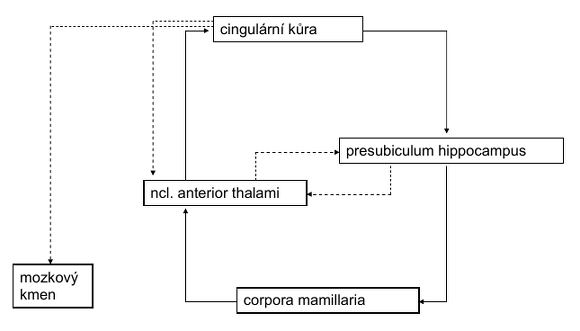
V limbickém systému není lokalizována žádná samostatná somato. –či viscero- motorická funkce, protože smyslové orgány nemají do tohoto systému přímý vstup. Přesto způsobují spoje kůry do thalamu a zpět a i asociační funkce korových oblastí smyslového vnímání, předávání získaných podnětů limbickým korovým bolastem a tím je současně každý smyslový vjem limbickým systémem vyhodnocen a zpracován.
Funkce limbického systému jsou složité a jeho výzkum není dokončen.
Funkce struktur limbického systému
Amygdalární systém. Účastní se na řízení vegetativních funkcí (starší pojmenování – viscerální mozek). Reguluje aktivity spojené se sexuálním chováním, projevy emocí a obživou. Dodává neutrálním vjemům emoční náboj. Vytváří paměť na zážitky založené jen na emocích, hlavně pokud byly spojeny s obavami. Vzpomeneme – li si později na tento zážitek, amygdala zpětně zpracuje vzpomínky, a my si prožijeme původní emoce. To nám umožňuje správně provádět obtížná a riskantní rozhodnutí v závislostech na vzpomínkách a na minulé emoční zkušenosti.
Zpracovává pocity ohrožení s následnou příslušnou sympatickou odpovědí. Umožňuje rozeznat výhružné výrazy tváře u ostatních lidí a určit směr upřeného dívání. Souborně je tedy corpus amygdaloideum pomocí svých spojení schopno integrovat senzorické informace ze zevních i vnitřních prostředí a na jejich podkladu ovlivňovat somatomotoriku a to pyramidové i mimopyramidové motorické systémy, jakož i visceromotorické reakce. Dráždění nebo neopak vyřazení amygdalární jáder vede ke změnám emoční aktivity a tím i ke změnám řady somatomotorických a zejména visceromotorických reakcí a ke změnám tvorby hormonů.
Porušení tohoto systému má za následek poruchu emočního prožívání. Vzniká Klüver-buckyho syndrom- destruovaného chování: nutkavé orální zkoumání jedlých i nejedlých předmětů, ztráta potravinových návyků, zánik agresivního chování, hyper- nebo asexualita, vizuální agnosie (neurologické porucha poznávání, ztráta schopnosti rozpoznat předměty, osoby, zvuky, tvary, vůně (bez porušení smyslových orgánů).
Čelenkový závit – gyrus cinguli. Umožňuje vnímat rozdíly mezi tím, co si myslíme a jak dovolíme vyniknout naším emocím pomocí gest. Přední část tohoto závitu rozpoznává bolest jako nepříjemný zážitek a řeší duševní rozpory v průběhu frustrující činnosti.
Hippokampální formace. Skladá se z hippocampu (mořský koník) a hippokampálního závitu. Hippokampální formace ukládají, slučují a pozdějí zpětně vyvolávají vzpomínky na události a fakta. Nejprve přijímá informace určené k zapamatování ze zbytku mozkové kůry, poté tato data zpracovává a vrací zpět do kůry, kde zůstávají uložena v dlouhodobé paměti.Poškození hippokampální formace vede k poruše krátkodobé paměti.
Limbický systém komunikuje s mnoha dalšími oblastmi mozku (hypothalamus, retikulární formace), což jsou oblasti,které ovládají naší orgánovou odpověď. Tím jsou s limbickým systémem spojeny i nadřazené řídící funkce dýchácí, srdeční činnosti, udržování tělesné teploty, regulace funkce trávícího systému, cév, žláz s vnitřní sekrecí a mnoha dalších funkcí autonomního nervového systému. Řada z těchto funkcí přímo souvisí s emočními reakcemi.. Tento fakt vysvětlujhe proč, lidé s emocionálním stresem trpí onemocněním jako je vysoký tlak nebo pálení žáhy. Limbický systém je také propojen s prefrontálními laloky mozkové kůry. Limbický systém řídí více biologickou složku lidské osobnosti, která je do jisté míry relativně autonomní, na rozdíl od neocortexu, sídla nejvyšších vrstev vlastního vědomí, řeči, abstraktního (tvořivého) myšlení, který je substrátem osobnosti člověka jako bytosti společenské. Tak jsou naše pocity (přenášené emočním mozkem) a naše myslenky (přenášené myslícím mozkem) těsně spjaty.
-
Hormony ovlivňující mozek
Pozn.: Pohlaví mozku aneb proč muži myslí tak, jak myslí a proč jednaji jinak než ženy, a proč ženy myslí tak, jak myslí a proč jednají jinak než muži. Regulace a řizení celého organismu je prováděno na několika úrovních – genetické, imunitní, humorální a nervové. Ale i tyto úrovně se navzájem významně ovlivňují, jednak přímo a jednak zpětnými vazbami. Rozdíl mezi mozkem muže a ženy je v genetické výbavě – a to především v chromozomech XX a XY. Dále je významný rozdíl od vývoje plodu a pak po celý život jedince v hormonální výbavě mezi pohlavími a jejím vlivem na činnost mozku. Činnost ženského i mužského mozku jsou významně ovlivněny změnami hormonů, zejména pohlavních.
Rozdíly fuknce mozku dle pohlaví
Oblast mozku
Ženy
Muži
Mesencephalon – střední mozek
Ventrální část tegmenta
Centrum motivace- oblast hluboko ve středu mozku, která produkuje dopamin, neurotransmiter potřebný k zahájení pohybu, motivace a pocit odměny. Méně aktivní
Centrum motivace – oblast hluboko ve středu mozku, která produkuje dopamin, neurotransmiter potřebný k zahájení pohybu, motivace a pocit odměny. Je aktivnější
Periakveduktální šeď středního mozku
Součást mozkového obvodu bolesti, jenž pomáhá kontrolovat bezděčné požitky a bolest. Během pohlavního styku je toto centrum odpovědné za potlačení bolesti, intenzivní požitky a sténání, Méně aktivní.
Součást mozkového obvodu bolesti, jenž pomáhá kontrolovat bezděčné požitky a bolest. Během pohlavního styku je toto centrum odpovědné za potlačení bolesti, intenzivní požitky a sténání. Během sexu je aktivnější
Mezimozek
Hypothalamus
Dirigent symfonie hormonů. Uvádí gonády do chodu. Začíná „pumpovat“ dříve u žen.
Dorsální premammilární jádro
Oblast obrany vlastního teritoria leží hluboko uvnitř hypothalamu a obsahuje systém obvodů zodpovědných za mužské instinkty silného vůdčího samce, obrany teritoria, strach a agrese. U žen malé.
Oblast obrany vlastního teritoria leží hluboko uvnitř hypothalamu a obsahuje systém obvodů zodpovědných ze mužské instinkty silného vůdčího samce, obrany teritoria, strach a agrese. Je větší u mužského než u ženského mozku a zahrnuje zvláštní okruhy k odhalování ohrožení teritoria dalšími muži, díky čemuž jsou muži citlivější na možné riziko ztráty svého území
Mediální preoptická oblast
Je to oblast sexuálního usilování, nachází se v hypothalamu, U ženské mozku je 2,5x menší než u mužského.
Je to oblast sexuálního usilování, nachází se v hypothalamu a u mužského mozku je dvaapůlkrát větší. Muži ji potřebují k zahájení erekce.
Podvěsek mozkový
Produkuje hormony plodnosti, tvorby mléka a mateřského pudu. Pomáhá mozku ženy změnit se na mozek matky.
Čelní laloku
Prefrontální kůra
Královna, která vládne nad emocemi a brání jim zdivočet. Tlumí činnost amygdaly. Je větší u žen a vyvíjí se u nich o 1–2 roky rychleji.
Je to generální ředitel mozku, soustěďuje se na aktuální dění a vytváří z něj správný úsudek.Tato oblast „absolutní pozornosti věnované na tady a teď“ je zároveň brzdným systémem, který drží na uzdě naše impulsy. Je větší v ženském mozku a dozrává asi o jeden až dva roky dříve u žen než u mužů.
Gyrus cinguli
Barometr mozku pro odhalování sociálního přijetí nebo odmítnutí. Tato oblast „jsem, či nejsem přijímán“ pomáhá lidem předcházet té nejzásadnější sociální chybě: příliš se lišit od ostatních. Gyrus cinguli je mozkové centrum zpracovávání společenských omylů. Dává nám varování, chováme-li se nepatřičně ve vztazích nebo v práci. Během puberty pomáhá chlapcům minimalizovat obličejové reakce, což umožňuje utajení jejich emocí.
Přední část cingulární kůry
Zvažuje možnosti, dělá rozhodnutí. Je centrum starostí a je větší u žen než u mužů.
Centrum starostí, strachu z potrestání a rovněž úzkosti o sexuální výkonnost. U mužů je menší než u žen. Zvažuje možnosti, odhaluje konflikty, motivuje rozhodování. Testosteron zvyšuje obavy z potrestání. Přední část singulární kůry je rovněž centrem nesmělosti a ostychu.
Spánkový lalok
Temporoparietální spoj
Mozkové centrum „kognitivní empatie „ je jakýsi hledač řešení – mobilizuje mozkové zdroje k řešení stresujících problémů v situacích, kdy je potřeba vzít v úvahu perspektivu jiné zúčastněné osoby nebo osob. Během mezilidských emočních výměn je méně aktivní u žen. Pomaleji se aktivuje. Řešení má vždy prodlevu.
Mozkové centrum „kognitivní empatie „ je jakýsi hledač řešení – mobilizuje mozkové zdroje k řešení stresujících problémů v situacích, kdy je potřeby vzít v úvahu perspektivu jiné zúčastněné osoby nebo osob. Během mezilidských emočních výměn je aktivnější v mužském mozku, rychleji se aktivuje a usiluje o co nejrychlejší řešení.
Insula
Centrum, které zpracovává emoce. Je větší a aktivnější v ženském mozku.
Centrum, které zpracovává emoce. U mužů menší.
Amygdala
Divoká šelma uvnitř mozku, jádro instinktů, zkrocené pouze prefrontální kůrou mozkovou. U žen menší až 2x.
Poplašný systém pro hrozby, strach a nebezpečí. Řídí emoční impulsy. Do stavu připravenosti k boji ji přivádí testosteron, vazopresin a kortizon, naopak její uklidnění způsobuje oxytocin. Tato oblast mozku je větší u mužů.
Hippocampus
Slon, který nikdy nezapomene hádku, romantickou chvilku nebo něžný moment – a nenechá vás nic z toho zapomenout. Je větší a aktivnější u žen.
Slon, který nikdy nezapomene hádku, romantickou chvilku nebo něžný momemt – a nenechá vás nic z toho zapomenout. Je menší a méně aktivní u mužů.
Systém zrcadlových neuronů
Systém emocionální empatie – „cítím to,co cítíš ty“. Umožňuje sladění s pocity druhých pomocí čtení výrazu jejich obličeje, interpretování barvy hlasu a dalších neverbálních emočních signálů. Je větší a aktivnější v ženském mozku
Systém emocionální empatie – „cítím to,co cítíš ty“. Umožňuje sladění s pocity druhých pomocí čtení výrazu jejich obličeje, interpretování barvy hlasu a dalších neverbálních emočních signálů. Méně vyvinuto a méně aktivní
Hormony ovlivňující mozek
Hormon
Muži
Ženy
Testosteron
Zeus. Král mužských hormonů: je dominatní, agresivní a všemocný. Je rozhodný a cílevědomí a zuřivě buduje všechno, co je mužské, včetně nutkání předčít všechny ostatní muže ve svém okolí. Pohání mužské potní žlázy k výrobě přitažlivého aromatu mužnosti – androstenedionu. Aktivuje obvody sexu a agrese a je naprosto cílevědomý ve své urputné honbě ze vytouženou družkou. Oceňovaný za svou smělost a statečnost, může se stát přesvědčivým svůdcem, ale je-li podrážděný, dokáže být tím nejmrzutějším neurvalcem.
Rychlý, důrazný, soustředěný, horlivý, mužský, silný svůdce, agresivní, necitlivý, nemá čas na mazlení.
Estrogen
Královna. Ačkoli nemá nad mužem takovou moc jako Zeus, dokáže být skutečnou ukrytou silou, neboť pohání většinu obvodů v mužském mozku. Má schopnost zvyšovat mužskou touhu po mazlení a vytváření vztahů jeho stimulováním tvorby oxytocinu.
Mocná královna, která vše ovládá, je horlivá, někdy ve všech oblastech, někdy je agresivní svůdnice, přítelkyně dopaminu, serotoninu, oxytocinu, acetylcholinu a norepinefrinu (chemické látky šťastných pocitů).
Progesteron
---
Méně nápadná, ale mocná sestra estrogenu. Občas se objeví, někdy jako bouřkový mrak, který převrací naruby účinky estrogenu, jindy jako zjemňující činitel. Je to matka allopregnenolonu (diazepam, tj. uklidňující lék pro mozek).
Vazopresin
Bílý rytíř. Vazopresin je hormonem rytířskosti a monogamie, jenž agresivně ochraňuje a brání své území, partnerku a děti. Spolu s testosteronem pohání mužský mozek a umocňuje mužskost.
Nenápadná mužská energie v pozadí, bratr testosteronu a oxytocinu) stejně jako oxytocin způsobuje, že se žena chce aktivně zapojovat – mužským způsobem).
Oxytocin
Krotitel lvů. Stačí jen několik málo objetí a pohlazení a tento“ konejšivý“ hormon utiší i tu nejzuřivější bestii. Zvyšuje schopnost empatie a buduje obvody důvěry, romantické lásky a silného pouta k druhému. Redukuje stresovoé hormony, snižuje tlak a hraje roli ve vytváření otcovského pouta k potomkům. Podporuje pocity bezpečí a jistoty a nese vinu ze mužskou „postkoitální narkolepsii“.
Chundelatá, vrnící kočička; pečující, hýčkající matka Země, dobrá čarodějka Linda z čaroděje ze země Oz, nachází potěchu v pomáhání a obsluhování; sestra vasopresinu (mužský socializační hormon), sestra estrogenu, přítelkyně dopaminu (další chemická látka šťastných pocitů). Vyvolává silnou touhu po dítěti.
Prolaktin
Pan Maminka. Způsobuje „sympatetické těhotenství“ (syndrom couvade) u nastávajících otců a zvyšuje schopnost tatínků slyšet pláč jejich dětí. Stimuluje spoje mužského mozku pro rodičovské chování a zvyšuje sexuální zaujetí.
Pan Maminka. Stimulátor mateřství a silného pouta k dítěti.
Androstenedion
Romeo.Okozlující svůdce žen. Uvolní-li ho pokožka v podobě feromonu, zvýší mužký sex-appeal víc než nejdražší kolínská.
---
Dehydroepiandrosteron (DHEA)
Rezervoár všech hormonů, všudypřítomný, vše prostupující, esence života dodávající energii, otec i matka testosteronu a estrogenu, přezdívaný „mateřský hormon“, Zeus a Héra všech hormonů, v mládí je přítomen v obrovském množství, ve stáří se jeho hladina snižuje, až vymizí docela.
Rezervoár všech hormonů, všudypřítomný, vše prostupující, esence života dodávající energii, otec i matka testosteronu a estrogenu, přezdívaný „mateřský hormon“, Zeus a Héra všech hormonů, v mládí je přítomen v obrovském množství, ve stáří se jeho hladina snižuje, až vymizí docela.
Allopregenolon
---
Skvělá, konejšívá, zjemňující sestra progesteronu, bez které jsme mrzuté,. Je uklidňující, zlehčující, neutralizuje stres, ale jakmile zmizí, všechno nám leze na nervy. Její odjezd tvoří hlavní děj příběhu premenstruačního syndromu, tří nebo čtyři dny před začátkem menstruace.
Dopamin
Životabudič. Tento opojný nápoj se stará o všechno spojené s příjemnými pocity, zábavou a uživáním života. Je rozvášněný, vysoce motivovaný a pudí muže k výhře, a znovu a znovu sázet na jackpot. Ale pozor – slast, kterou poskytuje, je návyková, zvláště v zápasnických hrách chlapectví a sexuálních hrách mužství, kde dopamin během orgasmu umocňuje pocity extáze.
Životabudič. Tento opojný nápoj se stará o všechno spojené s příjemnými pocity, zábavou a uživáním života. Je rozvášněný, vysoce motivovaný. Pocit slasti. Ale pozor – slast, kterou poskytuje, je návyková. Dopamin během orgasmu umocňuje pocity extáze.
Kortisol
Gladiátor. V ohrožení vzplane hněvem a je ochotný bojovat o život do roztrhání těla.
Prskající,vyčerpaný,, přepracovaný, přecitlivělý tělesně i emočně
Müllerianova inhibiční substance (MIS)
Herkules. Je silný, houževnatý a neuhrožený. Je rovněž známý jako „defeminizátor“, protože nemilosrdně vyhání z muže vše, co je ženské. MIS vytváří mozkové obvody objevitelského chování, potlačuje obvody pro chování ženského typu, ničí ženské reprodukční orgány, a naopak pomáhá vybudovat mužské rozmnožovací orgány a mozkové obvody.
-----
Hladiny hormonů se mění v průběhu života, ale i v průběhu měsíce a dne. Mají výrazný cyklický charakter.
Pokud se jedná o termíny puberta a menopauza – ty se v časové ose odehrají fyziologicky v průběhu 24 hodin. Stav před pubertou a po uplynutí 24 hodin již není vlastní fyziologický akt puberty. Rovněž u menopauzy – stav před je premenopausální období a po uplynutí 24 hodin pak postmenopausální období. Jsou to stavy, které s mozkem a jeho činnosti významně zamávají a změní jeho způsob činnosti. Např. pokud bychom u chlapců přirovnali hladinu testosteronu před pubertou k jednomu půllitru piva denně, pak v průběhu 24 hodin puberty dojde k jeho zvýšení, jako by místo jednoho půl litru piva vypil 7,5litru piva za jeden den.
Mužský se ženský mozek se o sebe liší již od chvíle početí. Mužská buňka obsahuje chromozom Y a ženská nikoli. Již osm týdnů po početí začínají maličké varlata produkovat dostatek testosteronu, aby jím zaplnila mozek a změnila od základů jeho strukturu.
V ženském mozku předurčují hormony estrogen, progesteron a oxytocin mozkové obvody k typicky ženskému chování. V mužském mozku má nejranější a nejdéle přetrvávající vliv testosteron, vazopresin a látka zvaná Müllerianova inhibiční substance (MIS). Např. muži používají zcela jiné mozkové obvody ke zpracování prostorových informací a řešení citových problémů. Mozkové obvody a nervový systém mají odlišné napojení na své svaly – zejména v obličeji. Muži mají dvaapůlkráte větší prostor v mozku, konkrétně v hypothalamu, zasvěcený sexuálnímu pudu. Mají i větší část mozku, která je zasvěcená agresi, připravenosti k boji, poplašný systém pro hrozbu, strach a nebezpečí, Mají rozsáhlejší mozková centra pro svalovou aktivitu a aktivnější oblasti pro ochranu partnerky a bránění svého teritoria.
Vývoj u ženy
Fáze
Významné hormonální změny
Rozdíl ženy muži
Specifické změny v ženském mozku
Změny vnímání reality
Fetální (plodová)
Růst a rozvoj mozku se vyhne vysoké hladině testosteronu vyskytující se v mozku mužském
Mozkové buňky jsou XX, což znamená více genů pro rychlý rozvoj mozku a okruhů specifických pro ženy
Mozkové okruhy pro komunikace, prožívání emocí, citovou paměť a potlačování hněvu neomezeně rostou – v okolí není žádný mužský testosteron, který by všechny tyto buňky zabíjel
Více mozkových okruhů pro komunikaci, „čtení“ v emocích, společenských nuancích, schopnost operovávat, schopnost používat obě hemisféry
Dívčí věk
Estrogen je v obrovském množství vylučován ve věku od šesti do dvaceti čtyř měsíců. Poté se až do puberty příliv hormonu přeruší.
Vysoká hladiny estrogenu do dvou let po narození.
Verbální a emoční okruhy se zvětšují
Hlavním zájmem je hra a legrace s ostatními děvčaty, ne s kluky
Puberta
Hladina estrogenu, progesteronu a testosteronu se zvyšuje a získává měsíční cyklus.
Více estrogenu a méně testosteronu; dívčí mozek začně dospívat o dva roky dříve než chlapecký
Zvyšuje se citlivost, narůstají verbální, emoční a sexuální okruhy.
Hlavní roli hraje sexuální přitažlivost, beznadějná zamilovanost, vyhýbání se rodičům.
Sexuální zralost, žena bez partnera
Hladina estrogenu, progresteronu a testosteronu se mění každý den v měsíci.
Více se soustředí na vztahy, na hledání životního partnera a kariéry či práce, která by se dala skloubit se založením rodiny.
Dřívější dospívání okruhů rozhodování a kontroly emocí.
Nejdůležitější pro ženu je najít lásku, životního partnera a budovat kariéru.
Těhotenství
Obrovský nárust hladiny progesteronu a estrogenu
Soustředí se více na péči o rodinu, méně na práci a soutěživost.
Stresové okruhy jsou potlačeny; mozek je zklidněn progesteronem; mozek se scvrkne; hormony z plodu a placenty převezmou velení nad tělem i mozkem.
Středem zájmů je vlastní tělesná pohoda, zvládání únavy, nevolnosti a hladu a snaha neublížit plodu, přežít v práci a plánovat mateřskou dovolenou.
Kojení
Oxytocin, prolaktin
Mnohem více se zaměřuje na dítě
Stresové okruhy jsou stále potlačeny, péče o dítě se zmocnila sexuálních a emočních okruhů.
Ženy myslí na to, aby zvládla únavu, bolest bradavek a produkci mateřského mléka i v příštích dvacetičtyřech hodinách.
Výchova dětí
Oxytocin a v měsíčních cyklech estrogen, progesteron a testosteron.
Méně zájmu o sex, více si dělá starosti o děti.
Vzrůstá aktivita oblastí spojených se stresem, obavami a emočními pouty.
Nejdůležitější je pohoda, rozvoj, vzdělání a bezpečnost dětí a zároveň zvládání narůstajícího stresu a práce.
Období před menopauzou
Nevyzpytatelně kolísající hladiny estrogenu, progesteronu a testosteronu.
Kolísavý zájem o sex, nepravidelný spánek, větší únava, obavy a utrápenost, návaly horka a podrážděnost.
V některých mozkových okruzích se snižuje citlivost na estrogen.
Cílem tohoto období je přežít ze dne na den a zvládat fyzické i citové výkyvy.
Menopauza
Nízká hladina estrogenu, žádný progesteron, vysoká hladiny FSH/LH
Poslední náhlá změna v mozku způsobená hormony.
Okruhy poháněné estrogenem,oxytocinem a progesteronem ochabují.
Hlavním zájmem je zůstat zdravá, zlepšovat pohodu a chopit se nových příležitostí.
Období po menopauze
Nízká hladiny estrogenu a testosteronu, nižší oxytocin
Více klidu
Okruhy jsou méně citlivé na stres, méně vzrušivé.
Nejdůležitější je dělat to, co chcete vy; méně zájmu pečovat o ostatní. Méně zájmu o sex.
Vývoj u muže
Fáze
Hlavní hormonální změny
Rozdíl muži ženy
Změny v mozku typické pro muže
Reálé změny v chování muže
Fetální – vývoj plodu
Vývoj mozku; 8 týdnů po početí začne testosteron maskulinizovat (pomužšťovat) mužský mozek, poté ho spolu s MIS defeminizuje (odženšťuje)
Chromozóm Y
Růst a maskulinizace center sexuality, objevitelského chování a svalových pohybů pro boj.
Chlapectví
Pokračuje tvorba MIS; během této „juvenilní pauzy“ je v těle nízká hladiny testosteronu.
Vysoká hladiny testosteronu od 1 do 12 měsíců po narození (juvenilní puberta); nižší hladiny testosteronu od 1 do 11 let; stále velké množství MIS; nízká hladina estrogenu.
Rozšiřování mozkových okruhů objevitelského chování, bojových svalových pohybů; okruhy mužské sexuální aktivity se stále rozvíjejí.
Hlavním zájmem je vyhrávání, pronásledování, boje a průzkumnické hry s chlapci, nikoliv s dívkami.
Puberta
Dvacetinásobné zvýšení hladiny testosteronu spolu s příbytkem vazopresinu; nízká hladiny MIS.
Zvýšená citlivost a růst center sexuálity a teritoriální agresivity.
Okruhy pro zrakovou sexuální přitažlivost se soustředují na ženské postavy; mužské tváře vnímá jako nepřátelské; mění se smysl pro čichové vnímání feromonů; mění se sluchové vnímání; mění se centra cyklů spánku.
Hlavním zájmem je vlastní teritorium, sociální interakce, části dívčího těla, sexuální fantazie, masturbace, hierarchie mezi muži; chodí později spát a později vstává, vyhýbá se rodičům, zpochybňuje autoritu.
Pohlavní dospělost, svobodný muž
Testosteron má stále vysokou hladinu a aktivuje centa hledání partnerky, sexuality, ochraňování, hierarchie a obrany úzení.
Zaměření na dobře stavěné, plodné ženy. Chce nejprve sex, láska a vztah mohou následovat později; silné libido.
Zrakové okruhy se mění k lepšímu rozeznání plodných žen a potenciálně agresivních mužů.
Hlavní zájem v hledání sexuálních partnerek; zaměření na práci, peníze a rozvoj kariéry.
Otcovství
Během těhotenství partnerky a po narození dítěte se zvyšuje množství prolaktinu a snižuje hladiny testosteronu.
„mužské těhotenství“ neboli syndrom couvade
Centra sexuálního pudu ustupují do pozadí díky nižší hladině testosteronu a většímu množství prolaktinu; sluchové okruhy jsou citlivější k vnímání dětského pláče; rozvíjí se, synchronie mezi otcem a dítětem.
Hlavním zájmem je ochrana matky a dítěte, zajištění živobytí a podpora rodiny; slyší děti plakat lépe než muži, kteří nejsou otci.
Střední věk
Velmi pozvolna klesají hladiny testosteronu
Přetrvává zaměření na sex, vlastní území a atraktivní ženy.
Zvolna se snižuje aktivace testosteronem a vazopresinem.
Hlavní zájmem je výchova dětí, moc a postavení v práci; menší zaměření na okamžitou potřebu sexu.
Andropauza
Postupně klesá hladiny testosteronu; kolem 85let je jeho množství méně než poloviční oproti věku 20 let.
Může pokračovat v reprodukci; přetrvává zaměření na sex a atraktivní ženy.
Mozkové obvody,dosud zásobované testosteronem a vazopresinem, začínají slábnout; poměr estrogenu vůči testosteronu se zvyšuje; vyšší hladina oxytocinu.
Hlavním zájmem je zůstat zdravý, zachovat si blahobyt a zlepšovat své manželství, sexuální život, mít vnoučata a vytvořit svůj odkaz; z celého živost jsou muži v tomto období nejvíce podobni ženám, protože oxytocin je činí otevřenějšími k náklonnosti a porozumění a snižující se hladiny testosteronu jim ubírá na agresivitě.
Mužský mozek vizuálně zkoumá pohybující se objekty zatímco ženský mozek pohyb, tedy mimiku, tváře. Ve vývoji chlapce je nacházení místa na světě dáno posouvání veškerých fyzických hranic svého těla, včetně hraní si s penisem, snaží se demonstrovat svou sílu. Tedy jedná se o budování fyzické a sociální dominance. Při učení u chlapců jsou více využívány svaly a nervový systém častějí než u dívek. Při výuce jsou proto více neposední. Vyjadřují se více pomocí pohybu. Od pěti let se objevuje velký vývoj v chápání prostoru a jeho vztahů u chlapců – jde o propojení vizuální kůry a parietálního centra prostového pohybu v obou hemisférách – oproti dívkám. V době dospívání se objevuje výrazný rys teritoriality a dále změny denního cyklu – chlapci se čtrnáci letech mají nově naprogramovanou dobu spánku – je posunutá asi o hodinu oproti stejně starým divkám. Dále dochází v této době k výraznému utlumení centra příjemných pocitů v mozku, dochází dále k výraznému omezení mimiky obličeje. Dochází k změnám ve sluchovém vnímání. – u mužských mozků je aktivace hudbou, ale nikoli tzv. bílým šumem (jednoduché zvuky, bílý šum je tak nazýván jako analogie s bílým světlem, které obsahuje všechny frekvence, je náhodný signál s rovnoměrnou výkonovou spektrální hustotou, signál má stejný výkon v jakémkoli pásmu shodné šířky) a opakujícími se akustickými stimuly, u žen obojím. Oblast mozku pro potlačování hněvu, je menší u muží než žen, a tak vyjadřování hněvu je obvyklejší odpovědí pro muže než pro ženy. Aby muž fyzicky zvládnul pozici sebedůvěry a síly, musí trénovat své obličejové svaly k zamaskování obav – tedy tzv. mužský výraz hráčů pokru.
Naše pohlavní hormony mohou být ve skutečnosti částečně zopovědné ze naše odlišné emocionální styly. Mužské obvody využívají víc testosteronu a vazopresinu; ženské zase pohání estrogen a oxytocin. Tyto hormony spouštějí jisté oblasti mozku (amygdala, hypothalamus, systém zrcadlových neuronů a temporoparietální spoje) u mužů odlišně než u žen. Chování jako blafování, předstírání nebo boj se vyvinulo proto, aby chránilo samce zejména před protivníky z jejich vlastního druhu. V sociálních interakcích dochází k změnám hladin hormonů a to ovlivňuje i způsob činnosti mozku (např. když muž vidí atraktivní ženu, z pohledu plodnosti, zvýší se mu hladina testosteronu, při dosažení určitého stupně v hierarchii společnosti dojde ke snížení hladiny testosteronu, kortizonu apod.). Hladinu obvlivňuje i fyzická zátěž – cvičení a přetrvávájcí sexuální aktivita.
Ženský mozek je silně ovlivněn hormony, že se o nich může říci, že vytváření ženskou realitu. Tato se mění cyklicky dle hormonálních změn. Příkladem je tzv. premenstruční mozkový syndrom, kdy u extrémních forem dochází k velké míře depresí u žen. U všech menstruujících žen probíhají malé změny v mozku každý den. Některé části mozku se změní každý měsíc o 25 %. Muži s ženy mají různě citlivý mozek na stres a konflikt. Ženy mají ve sluchovém a řečovém centru o11 % neuronů více než muži. Rovněž mají větší hipocampus. V průměru ženy jsou lepší se vyjadřování pocitů a snáze si pamatují detaily citově vypjatých událostí. Oproti tomu, muži mají v mozku dvaapůlkrát více prostoru zaměřeného na sexuální touhu a stejně tak mají větší mozková centra agrese a akce. Ženy mají vynikající schopnosti verbální, schopnost vytvořit hluboká přítelství, skoro okultní nadání vyčíst z tváří a tónu hlasu pocity a stavy mysli a dokáží zmírňovat konflikty. Muži řeknou asi sedm tisíc slov za den, ženy jich vysloví asi dvacet tisíc. Nárust hladiny testosteronu v osmém týdnu těhotenství zabije některé buňky v centru komunikace a vytvoří více buněk v centru agrese a akce. Během prvních 3 měsíců života se děvčátka zdokonalují v očním kontaktu a schopnost vzájemného pozorovní u nich vzroste o 400 %, zatímco u chlapců se tato schopnost nezdokonalí vůbec. Děvčátka se rodí se zájmem o vyjadřování emocí. Vytvářejí si o sobě mínění z pohledů, doteků, z každé reakce lidí, se kterými se dostanou do kontaktu. Jednoletí chlapci, řízení mužským mozkem utvářeným testosteronem, mají nutkání prozkoumávat své okolí, dokonce i ty jeho součásti, kterých se nesmějí dotýkat. Dívka je velmi vnímavá, co se týče čtení výrazu tváře, ať je jí nasloucháno nebo ne. Jednoletá děvčata více reagují na rozrušení ostatních lidí, zvláště těch, kteří vypadají nešťastně nebo ublíženě. Dívky jsou naprogramovány, aby se neustále ujišťovaly, že žijí s ostatními v souladu. Chlapci také vědí jak použít spolupracující styl řeči, ale obvykle jej nepoužívají. Obecně používají řeč, aby druhé ovládli, něco vyřídily, chvastali se, zastrašili, ignorovali návrhy nebo přešli pokusy ostatních něco říct.
Dovednost mužského a ženského mozku navázat mezilidské vztahy je významně ovlivněno nejen geny, ale množstvím testosteronu, které se do mozku embrya dostane. Podle všech měřítek jsou muži průměrně dvacetkrát agresivnější než ženy. Během puberty u dívky je smyslem celé existence být sexuálně přitažlivá. Je to dáno hormonálními změnami. Díky pubertě se objeví změny v rámci estrogen-progesteronové vlny. Zvýší se emocionální citlivost na okolí, zvýší se citlivost na stres. Dívku stresují konflikty ve vztazích. Potřebuje, aby ji měli rádi a udržovala společenské vztahy; dospivající chlapci potřebují být respektování a mít vyšší místo na dominančním řebřičku (tzv. klonovací řád).
Výsledky experimentů dále ukázaly, že chlapci mají více nadání na orientaci v prostoru, na rozlišování a skládání geometrických forem, zatímco děvčata obratně a výborně zarábějí se slovem. Na řešení stejného úkolu používají odlišnou techniku. Muži se ve městě orientují tak, že si osvojí celkový plán, ale ženy si určí orientační body, např. semafor nebo obchod. (Říkejme tomu ženský smysl pro detail.) Každá hemisféra má svou specifickou funkci. U muže je rozdělení mezi levou a pravou polokoulí jasně stanovené. Vpravo se nachází vnímání prostoru a forem, vlevo řeč a myšlení. Muž se tedy orientuje pravou polokoulí a učí číst levou. Ženy, či už jde o čtení, mluvení anebo kreslení plánů, používají obě mozkové polokoule.14
14 Hanzlovský M., Vojna pohlaví – ženský mozek versus mužský [online], www.celostnimedicina.cz, Dostupné z WWW: http://www.celostnimedicina.cz/vojna-pohlavi--zensky-mozek-versus-muzsky.htm
První dva týdny cyklu, kdy je hladina estrogenu vysoká, je dívka společenštější a uvolněnější. Poslední dva týdny, kdy jse hladině progesteronu vysoká a estrogen je nižší, má sklon reagovat více podrážděně a chtít být o samotě. Estrogen a progesteron nastaví citlivost na zátěž každý měsíc. Dívčino sebevědomí může být jeden týden vysoké a následující křehké. Pokud v době ovulace dojde k oplodnění vajíčka se situace zcela jiná. Spousta žen nachází biologickou podporu ve společnosti jiné ženy a mluvení je pojítko, které je stmeluje. Hladina estrogenu změní mozek tak, aby dívky více mluvily a komunikovaly s vrstevnicemi, myslely více na kluky, dělaly si starosti o svůj vzhled, byly citlivější a nervoznější. Způsobuje to příval dopaminu a oxytocinu. Chlapci nesdílí silnou touhu po verbálním kontaktu. Zvýšení hladiny testosteronu u mladého muže v poberte způsobí to, že se „vytratí dospívat“ – uzavře se ve svém teritoriu, které chrání a omezuje komunikaci, socializaci – s výjimkou sportu (pohyb, svalová činnost, boj i hierarchii v mužské společnosti) a snahou o sex.
Ženský mozek má daleko negativnější poplašní reakce než mužský. Muži si mezilidské spory a soupeření často užívají, dokonce je to posílí. Mužská hrdost pochází více ze schopnosti udržovat si nezávislost, zatímco ženská ze schopnosti udržovat důvěrní vztahy s ostatními.
Mozek ženy v podstatě skáče jak estrogen píská. Zvýšená hladiny estrogenu ovládá prakticky cokoli, reakce na světlo a denní režim světla a tmy. Estrogenové receptory se zaktivizují na buňkách, které fungují jako naše „vnitřní hodiny“ a jsou umístěny v suprachiasmatickém jádru. Tyto shluky buněk řídí denní, měsíční a roční rytmy našeho těla, tak jako cykly hormonů, tělesné teploty, spánku a nálady. Estrogen dokonce přímo ovlivňuje mozkové buňky, které řídí dýchání. Spouští specifický ženský cyklus spánku, stejně jako tvorbu růstového hormonu. Od puberty řídí estrogen časové nastavení všeho, co se odehrává v ženském mozku – ženský a mužský mozek začínají pochodovat v jiném rytmu. Ženský mozek dospívá v průměru o dva až tří roky dříve než mužský. Např. tendence žen jít spát a budit se trochu dříve než muži je rozdíl, který bude trvat až do menopauzy. Estrogen podporuje tvorbu spojů v hipocampu – asi 25 % spojů se vytvoří během prvních dvou týdnů cyklu.V této fázi je mozek bystřejší, více si pamatuje, myslí svižněji a rychleji. Po ovulaci začíná produkce progesteronu, který působí na nově vzniklé spoje v hipocampu jako herbicid – mozek se nejprve zklidní, pak začně být podrážděnější, méně soustředěný a zpomalenější. V posledních dnech cyklu progesteron prudce klesne, jako zklidňující účinek neočekávaně ustoupí a zanechá mozek chvilkově zmatený, přetížený a podrážděný. V této době se ženy snáže rozpláčou, jsou často mrzuté, přetažené, agresivní, oponující, zaujaté, nebo mají dokonce beznadějě a deprese. Ženy, jejich vaječníky produkují nejvíce progesteronu a estrogenu, jsou vůči stresu nejodolnější, protože mají více buněk produkujících serotonin (chemická látka, která způsobuje, že se cítíme uvolněně). Ty ženy, které mají estrogenu a progesteronu nejméně, jsou na zátěž citlivější, neboť mají méně buněk produkujících serotonin. Je to asi 10 % žen – ty mají tzv. premestruační dysforickou poruchu, která je dána nedostatkem serotoninu. Hormony, které jsou u mužů i žen spojovány a agresivitou se nazývají androgeny. Jejich hladina se začíná na začátku poberty a jejich produkce je na vrcholu v devatenácti letech u žen, a v jednadvaceti letech u mužů. Tří nejdůležitější androgeny v ženském těle jsou testosteron, DHEA(dehydroepiandrosteron) a androstenedion. Hormony pouze zvyšují pravděpodobnost, že se v určitých situacích chování projeví. Androgeny způsobují nejen agresivitu, ale i sexuální touhu. Zamilovanost, tzv milostní pouto způsobuje dopamin a oxytocin. Oxytocin v mozku přirozeně stoupně po 20vteřinovém objetí partnerem. Pro ženy je důležité zvýšení hladiny dopaminu a oxytocinu. U ženy se při sexuální spojení zvýší hladiny oxytocinu, u muže hladiny vasopresinu. Tyto hormony pak zvýší hladinu dopaminu v mozku.Oxytocin u obou pohlaví přináší uvolnění, pocit bezstarostnosti, sepětí a vzájemné spokojenosti. Muž potřebují doteky 2–3x častěji než ženy, aby si udrželi stejnou hladinu oxytocinu. Stres má rovněž rozdílný vliv u muže i ženy – u muže „nakopne“ milostné okruhy – jsou to vojáci, kteří si pod stresem z války přivedou domů nevěstu, u ženy má efekt opačný. Stresový hormon kortizon brání v ženském mozku činnosti oxytocinu, což náhle utne ženskou touhu po sexu a fyzickém doteku.
Stejně jako je v určitém okamžitu mužský mozek pod poklopcem, tak i ženský je pod sukní (penis, klitoris). Ženský orgasmus je důležitý pro to, aby se spermie dostaly do blízkosti vajíčka., a tím došlo k oplodnění. V době před ovulací se mění i mozkové okruhy čichové u žen, zejména na nepředvídatelné účinky mužských feromonů. Jsou – li ženy před ovulací (tzn. na vrcholu jejich měsíční plodnosti) vystaveny feromonům z mužských potních žláz nazývaných androstadienon (blízky příbuzný androstenedionu, nejvýznamnější androgen tvořený vaječníky), během šesti minut se jejich nálada vyjasní a soustředění se zostří. Tyto feromony nesoucí se vzduchem udrží ženu v dobré náladě ještě hodiny poté. Protože ženin orgasmus nasaje spermie hluboko do reprodukčních orgánů, při vyvrcholení se svůdným mužem je větší pravděpodobnost, že se spermie dostanou k vajičku. Tato vzrůstající možnost otěhotnět s partnerem, který je sexy – má lepší genovou výbavu – může být důvodem, proč jsou ženy standartně více přitahovány atraktivními muži ve druhém týdnu menstruačního cyklu, právě před ovulací (což je jejich nejkoketnější období v měsíc). V mužské i ženském těle je testosteron chemickým palivem, které uvádí do chodu sexuální motor mozku. Mateřství ženu změní, protože doslova přestaví její mozek. Jedná se o hormony z placenty – progesteron stoupá, mozek se zklidní, rozvolní se mozkové okruhy. Mozek těhotné ženy mezi šestým měsícem a koncem těhotenství se zmenšuje a přestavuje – některé části se zmenší, některé se zvětší. Úsudková část mozku se během těhotenství zvětší. V posledním jednom nebo dvou týdnech před porodem se mozek začně zvětšovat buduje rozsáhlou síť mateřských okruhů. Ochranitelský, mateřský mozek se soustředí na schopnost partnera zaopatřit rodinu, svými specifickými agresivními, ochranitelskými okruhy začně chránit své hnízdo a bezpečnost svých dětí. Matky mějí lepší prostorovou paměť než ženy, které ještě nerodily, a jsou přizpůsobivější a odvážnější, mají méně aktivní centra strachu. U mužů v posledních týdnech před porodem jejich partnerek se zvýší hladina prolaktinu, hormonu pro vylučování mateřského mléka, o 20 %. Dojde k dvojnásobnému zvýšení hladiny kortizonu, který zvyšuje vnímavost a ostražitost. V prvních týdnech po porodu klesne hladina testosteronu na třetinu, zatímco hladina estrogenu je vyšší než obvykle. Muži s nižší hladinou testosteronu dokážou lépe rozpoznat pláč svého dítěte, ale snižuje mužskou sexuální energii. Testosteron potlačuje mateřské chování u žen, i u mužů. U syndromu couvade – sympatické těhotenství, podobné jako u žen tzv. falešné těhotenství – mají muži vyšší hladinu prolaktinu, prudší pokles testosteronu – pravděpodobně tyto chemické změny mohou způsobovat feromony, které ženy vylučují během těhotenství a mění tak tajně prostřednictvím čichu svého milence na milujícího otce. Při aktu kojení jsou matka a dítě klíčový partneři. Hladina hormonů zvýšená kojením a kontakt tělo na tělo urychlují vytváření mateřských mozkových spojů. Čím déle a častejí dítě kojíme, tím větší je odezva prolaktinu a oxytocinu v mozku matky. Děti, kterým se dostalo mateřské péče méně se v dospělosti snáže dostanou do stresu, jsou hyperaktivnější, nepozornější, bojácnější a častěji nemocné. V nepředvídatelném prostředí začnou být matky ustrašené, plaché a děti vykazují známky deprese. Vytvořit předvídatelné prostředí je úkolem muže. Změny, které proběhnou v mozku matky, jsou nejzávažnější a nejtrvalejší v ženském životě. Mnoho evolučních psychologů se domnívá, že schopnost cítit bolest někoho jiného a rychle vyčíst výkyvy nálady umožnila pravěkým ženám pohotově vytušit potenciální nebezpečí nebo agresivní chování, vyhnout se následkům a ochránit tak sebe i své děti. Toto nadání také vybavilo ženy schopností odhadnout fyzické potřeby jejich dětí, které ještě neumí mluvit.
V mužském mozku vyvolávají emoce méně pocitů a více racionálních myšlenek. Hledá rychlé řešení.
Ženské slzy mohou v mužském mozku vyvolat bolest. Mužský mozek zaznamenává v bolestném výrazu bezmoc a je to pro něj těžké snášet. Ženy používají na citové zážitky obě mozkové hemisféry, zatímco muži jen jednu. U obou pohlaví je citovým strážníkem emocí amygdala, mandlový útvar, který se nachází ve spánkovém laloku. Amygdala je jako bezpečnostní alarm a koordinační systém, který zapojuje zbytek těla – střeva, kůži, srdce, svaly, oči, obličej, uši a nadledvinky – aby vyhlížely citové podněty. První stanicí emocí na cestě z amygdaly do těla je hypothalamus. Jako hlavní dispečink je zodpovědný za koordinaci systémů, které zvýší krevní tlak, srdeční tep, rychlost dechu a stimuluje reakce „útěk, nebo boj“ po obdržení zpráv z těla. Amygdala také varuje mozkovou kůru, „zpravodajský odbor“, který posoudí citovou situaci, analyzuje ji a určí, kolik pozornosti si zaslouží. To je okamžik, kdy jsme zaplavení vědomými pocity. Do této chvíle se odehrávají všechny mozkové procesy mimo smysly. Centrum pro rozhodování nebo výkonná složka – prefrontální kůra – se nyní rozhodne, jak zareagovat. Ženy mají relativně větší hipocampus, mají lepší paměť pro detaily, jak příjemné, tak nepříjemné, a to v jakékoli situaci. U mužů pouze pokud je amygdala aktivována ohrožením vztahu nebo fyzickým nebezpečím.
Další rozdíl mezi mužský a ženským mozkem je ve zpracovávání hněvu. Ačkoli muži i ženy odpovídají stejným množstvím zloby, vyjádření vzteku a agrese je jasně větší u mužů. Amygdala je centrem mozku pro zpracování strachu, hněvu a agrese a je fyzicky větší u mužů, zatímco centrum kontroly těchto pocitů – prefrontální kůra – je relativně větší u žen a rovněž i oblast septa. Amygdala má hojně receptorů na testosteron. Ženy často namísto vzteku spustí verbální okruhy. Ženy, když jsou na někoho naštvané, nejdříve o tom mluví s druhými. Po pomalejší reakci, pak rozjedou své rychlejší verbální okruhy, se kterými nemůže muž soupeřit. Průměrný muž mluví méně a jeho řeč není tak plynulá jako u žen. Mužské mozkové okruhy a tělo se pak můžou přepnout do fyzického vyjádření hněvu.Citvoé okruhy pro hněv a pocit bezpečí se v ženském a mužském mozku liší. Muži a ženy mají jiné emocionální nastavení mozku pro pocity strachu a bezpečí, posílené konkrétními životními zkušenostmi. Pocit bezpečí je zabudován v mozkových spojích. Ženský mozek se aktivuje více než mužský, pokud ženy očekávají strach nebo bolest.
Úzkost je stav, který se objeví, když stres nebo strach spustí činnost amygdaly, což způsobí, že mozek vědomě soustředí všechnu svou pozornost na blízkou hrozbu. Úzkost je 4x běžnější u žen. V době menopauzy žena zahazuje všechno a všechny a začíná znovu, od začátku.žena v přechodu si už nedělá takovou hlavu s tím, jak potěšit své okolí. Teď chce pro změnu potěšit sebe. S nízkou hladinou estrogenu se snižuje i hladina oxytocinu. Žena se méně zajímá o drobné výkyvy v emocích, méně se soustředí na udržování pohody a nedostává“dopaminovou horečku“ z některých věcí, tak jako dříve (dokonce ani z klábosení s kamarádkami). Nedostává uklidňující oxytocin odměnou za péči a starost o své ratolesti, proto věnuje menší pozornost potřebám jiných. V menopauze dojde ke snížení hladiny estrogenu, ale i testosteronu, a tím se změní i chuť na sex. Více než 65 % rozvodů u dvojic po padesátce je iniciováno ženami. Mužský a ženský mozek stárnou různou rychlostí, přičemž muži ztrácení kůru dříve než ženy. Jediné období, kdy má žena ustálené reakce na zátěž díky stálé a nízké hladině hormonů je v dětském období klidu nebo během těhotenství, kdy jsou buňky hypothalamu vyřazeny z chodu a citlivost na stres je nízká.
Primárně je mužský mozek objemově i povrchovou plochou až o 11 % větší než u žen. S věkem ale dochází k rychlejší ztrátě objemu i plochy u mužů než u žen. To může být i v důsledku termodynamických zákonů.
Z pohledu biologické antropologie jsou na základě hormonů a uvolnění neurochemických látek do organismu vytvořeny čtyři různé temperamenty (osobnosti), které vysvětlují, proč jsou lidé přitahováni k ostatním.
- Průzkumník má vysokou hladinu dopaminu, hledá dobrodružství a novost, je zvědavý.
- Tvůrce vykazuje vysokou aktivitu serotoninu, je klidný, má rád plánování a řády, je konvenční.
- Ředitel je ovlivněn testosteronem, myslí logicky, analyzuje.
- Vyjednavač má vysokou estrogenní aktivitu, je soucitný a nápaditý.
Pozn:
Koncový mozek – telencephalon
U člověka tvoří největší část koncový mozek (telencephalon). Je pokryt mozkovými rýhami (sulci cerebrales) a závity (gyri cerebrales). Větší rýhy oddělují 5 telencephalických laloků:
- frontální (čelní)
- parietální (temenní)
- okcipitální (týlní)
- temporální (spánkový)
- insulární (ostrovní)
↓ rozbalit celou poznámku ↓Pozn:
Koncový mozek – telencephalon
U člověka tvoří největší část koncový mozek (telencephalon). Je pokryt mozkovými rýhami (sulci cerebrales) a závity (gyri cerebrales). Větší rýhy oddělují 5 telencephalických laloků:
- frontální (čelní)
- parietální (temenní)
- okcipitální (týlní)
- temporální (spánkový)
- insulární (ostrovní)
Koncový mozek je nejrostrálnější a objemově největší částí mozku. Za vývoje má dvě části. Nepárovou část (telencephalon medium), která v definitivním stavu ohraničuje část III. mozkové komory před foramen interventiculare a je ukončená lamina terminalis, a párovou část která se rozvíjí do definitivní podoby mozkových hemisfér. Hemisféry se při intenzivním růstu podkovovitě stáčejí, což se odráží i na tvaru na ncl. caudatus, postranních komor, corpus callosum, fornixu. Na hemisférách rozlišujeme část pokrývající ostatní struktury – plášť – pallium a část rozvíjející se uvnitř hemisfér – bazální ganglia. Pallium podle fylogenetického vývoje rozdělujeme na paleopallium rozvíjející se v souvislosti s čichovou aferentací, archipallium, které je u člověka redukováno na dorsomediální plochu hemisféry a vývojově nejmladší neopallium, které u člověka dosahuje největšího rozvoje. Cytoarchitektonicky se paleopallium a archipallium skládají ze tří vrstev – allocortex a neopallium obsahuje šest vrstev uspořádaných v celém rozsahu obdobně – isocortex. Člověk patří k savcům gyrencefalním tzn., že povrch telencephala je zprohýbán do mozkových závitů – gyrů. Míra gyrifikace je u savců závislá na vzájemném poměru velikosti povrchu těla a velikosti lebky.
Charakteristickým rysem povrchu koncového mozku jsou mozkové závity – gyri, přibližně jeden cm široké, oddělené rýhami – sulci.
Výsledný tvar koncového mozku je určen masivním rozvojem neocortexu, který zatlačí palecortex na dolní a vnitřní plochu hemisféry (asi 1 % povrchu) a archikortex na vnitřní plochu hemisféry (asi 3,5 % povrchu). Mozkové hemisféry jsou vyklenuté na superolaterální ploše, na spodní a mediální oploštělé, mimo spánkového laloku,který je mírně vyklenutý. Čelní laloky jsou uloženy v přední jámě lebeční, spánkové ve střední jámě lebeční a týlní laloky nad tentorium cerebelli,v zadní jámě lebeční, jámy mozečkové – ve fossae cerebrales.
Facies superolateralis – horní, zevní plocha hemisféry.
Facies medialis – vnitřní plocha hemisféry, celá je přehlédnutelná až po odstranění mozkového kmene.
Facies inferior – spodní plocha hemisféry, v zadní části přechází do mediální plochy.
Margo superior – přechod facies superolateralis na facies medialis.
Margo inferomedialis – vnitřní okraj, kterým přechází facies inferior ve facies medialis. Je patrný pouze v přední části hemisféry.
Margo inferolateralis – dolní zevní okraj, kerým přechází facies superolateralis do facies inferior.
Polus temporalis – nejventrálnější místo temporálního laloku
Polus frontalis – nejventrálnější místo frontálního laloku.
Polus occipitalis – nejdorzálnější místo occipitálního laloku.
Lobus frontalis – část mozkové hemisféry mezi frontálním pólem a sulcus centralis.
Lobus parietalis – část mozkové hemisféry mezi sulcus centralis a sulcus parietooccipitalis.
Lobus occipitalis – část mozkové hemisféry mezi sulcus parietooccipitalis na mediální straně hemisféry, incisura preoccipitalis na margo inferolateralis a occipitálním pólem.
Lobus temporalis – část mozkové hemisféry pod sulcus lateralis a před incisura preoccipitalis.
Insula, lobus insularis – část hemisféry, která při vývoji hemisfér roste pomaleji a zůstává uprostřed zevní plochy hemisféry skryta operculy.
Sulcus lateralis (Sylvii) – hluboká rýha mezi čelním a spánkovým lalokem v přední části, mezi temenním a spánkovým v zadním úseku. V hloubce sulcus lateralis je fossa lateralis jejíž dno tvoří insula.
Sulcus lateralis – ramus posterior – zadní část laterální rýhy, kolem níž se zatáčí gyrus supramarginalis.
Sulcus lateralis – ramus ascendens je rýha, která odděluje pars triangularis a pars opercularis gyrus frontalis inferior.
Sulcus lateralis – ramus anterior je rýha, která odděluje pars triangularis a pars orbitalis gyrus frontalis inferior.
Operculum frontale – část čelního laloku překrývající insulu.
Operculum parietale – část lobulus parietalis inferior překrývající insulu.
Operculum temporale – část gyrus temporalis superior překrývající insulu.
Fossa lateralis cerebri – prostor, který je hlubokým pokračováním sulcus lateralis, jeho dno tvoří insula.
Sulcus centralis (Rolandi) – rýha oddělující lobus frontalis od lobus parietalis na zevní ploše hemisféry. Obvykle zasahuje i na mediální plochu.
Incisura praeoccipitalis – otisk laterální části margo superior pyramidy kosti spánkové na margo inferolateralis hemisféry. Tvoří hranici mezi spánkovým a týlním lalokem.
Sulcus parietooccipitalis – rýha mezi lobus parietalis a occipitalis na mediální ploše hemisféry. Kraniálně zasahuje na margo superior.
Fissura longitudinalis cerebri – odděluje v sagitální rovině obě mozkové hemisféry. Zasahuje sem duplikatura tvrdé pleny – falx cerebri.
Fissura transversa cerebri – (fissura telodiencephalica) – prostor nad volným povrchem thalamů a stropem III.mozkové komory, pod fornixy, navnitř od zevního povrchu je tela choroidea postranních komor
Neokortex je složen ze šesti vrstev buněk. Pro obdobné uspořádání a vývoj ve všech jeho částech mluvíme o isokortexu. Přesto neokortex vykazuje okrskové rozdíly v neuronálním uspořádání mozkové kůry. Jednotlivé okrsky neodpovídají jednotlivým závitům. Sledováním těchto rozdílů se zabývá architektonika mozkové kůry. Mluvíme o členění cytoarchitektonickém, myeloarchitektonickém, glioarchitektonickém, synaptoarchitektonickém, myelogenetickém a angioarchitektonickém. Podle architektoniky jsou vytvářeny "mapy" mozkové kůry. Nejpoužívanější, opakovaně ověřená je Brodmanova cytoarchitektonická mapa. Rozděluje isokortex na 11 krajin a 52 okrsků (areae). Rozdíly ve stavbě kůry souvisí s jejím funkčním zapojením. Rozlišujeme primární korové oblasti (projekční) s přesným topickým uspořádáním zapojení, sekundární korové oblasti, jejichž topické zapojení je méně detailní. Sekundární oblasti můžeme rozdělit na suplementární a asociační.
Primární motorická korová oblast (area 4) - oblast gyrus precentralis a přední části gyrus paracentralis. Odtud vychází podněty k vedoucí k pohybům svalů opačné poloviny těla. Somatotopické uspořádání je zobrazováno jako homunkulus. Na mediální straně hemisféry oblast pro svaly nohy a bérce, na horním okraji hemisféry pro svaly pletence pánevního, na zevní straně pro trup a hlavu. Největší rozsah mají oblasti pro svaly ruky, obličeje a svaly okohybné. Čím jemnější a přesnější pohyby svaly vykonávají, tím větší plochu zaujímá jejich korová oblast. Aferentní vlákna přichází z anterolaterálních jader thalamu a mozečku, eferentní vlákna jsou součástí tractus corticospinalis a corticonuclearis.
Premotorická korová oblast (area 6) – oblast před primární motorickou kůrou. Je zapojena do okruhu striatum – palidum – thalamus – kůra. Z oblasti vycházejí dráhy do ncl. ruber, do RF prodloužené míchy i do míchy a motorických jader kmene. Oblast je aktivní při přípravě složitých volních pohybů.
Suplementární motorická korová oblast - zaujímá část premotorické korové oblasti na mediální ploše hemisféry, je považována za místo, kde vzniká podnět k pohybu, poškození oblasti vede k zástavě pohybů i řeči.
Frontální okohybné pole (area 8 a 9) - oblast leží v zadní části gyrus frontalis medius. Přijímá vlákna z primární i sekundární zrakové oblasti, odvodná vlákna běží do area pretectalis a do colliculus superior. Oblast je odpovědná za koordinaci pohybů očí, hlavy a krku.
Brocovo motorické centrum řeči - v zadní části gyrus frontalis inferior (area 44, 45), a zadní části gyrus frontalis medius (část area 9). Aferentní vlákna přichází ze senzitivních oblastí kůry a z thalamu. Odvodná vlákna běží k motorickým jádrům svalů mluvidel (area 44, 45) a svalů ruky zůčastněných při psaní (area 9). Porucha centra vede k poruše vyjadřování mluvenou řečí a poruše psaní.
Prefrontální korová oblast je větší oblast před premotorickou korovou oblastí. Zahrnuje gyri frontales superior, medius et inferior, přední část gyrus cinguli. Má reciproční spoje s ostatními korovými oblastmi, thalamem, hypothalamem, corpus striatum, s druhostranným prefronálním polem. Eferentní vlákna jdou také do tractus frontopontinus. Tato oblast ovlivňuje individuální osobnostní rysy, intenzitu vnímání, schopnost učení a koncentrace, vyjadřování emocí.
Primární senzitivní korová oblast (area 3, 2, 1) - oblast gyrus postcentralis až na mediální stranu hemisféry. Oblast je přesně somatotopicky uspořádána. Senzitivní homunkulus je uložen obdobně jako motorický, největší plochu zaujímají části těla s největší citlivostí. Do oblati vstupují thalamokortikální dráhy přepojující senzitivní podněty přicházející cestou lemniscus medialis, tractus spinothalamicus a trigeminothalamicus. Dále sem přicházejí spoje z motorické korové oblasti. Eferentní vlákna spojují oblast s primární motorickou oblastí, motorickými kmenovými a míšními centry. Oblast zajišťuje uvědomování si doteku, diskriminace, polohy a pohybu těla.
Sekundární senzitivní korová oblast (area 5, 7) - je uložena za primární senzitivní oblastí v parietálním laloku. Aferentace přichází z thalamu a z primární motorické kůry. Odstředivá vlákna spojují oblast s primární senzitivní i motorickou oblastí. Centrum se podílí na komplexním vnímání senzitivních podnětů včetně bolesti, integruje vjemy povrchové i hluboké, uplatňuje se při prostorové paměti a při vnímání pohybu těla. Při poškození oblasti vzniká taktilní agnosie – postižený nedokáže hmatem rozpoznat předměty, má poruchy prostorové hmatové orientace.
Primární zraková korová oblast (area 17) – oblast v sulcus calcarinus a podél něj, část zasahuje na zevní plochu hemisféry. Oblast je retinotopicky uspořádána, střídají se okrsky přijímající podněty z pravého a levého oka. Do oblasti vstupují cestou radiatio optica vlákna z corpus geniculatum laterale – ze stejnostranných polovin sítnic obou očí. Vzruchy z levých polovin sítnic (pravá zorná pole obou očí) konči v levém corpus geniculatum a z pravých polovin sítnic v pravém corpus geniculatum laterale. V levé hemisféře tedy končí vzruchy z levých polovin sítnic obou očí. Vzruchy z horních polovin sítnic končí nad sulcus calcarinus, vzruchy z dolních polovin sítnic pod sulcus calcarinus. Odstředivá vlákna primární zrakové kůry jdou do sekundární zrakové kůry, do area pretectalis, colliculi superiores a ncll. pontis. Centrum je odpovědné za rozeznávaní tvaru, barvy a pohybu předmětů.
Sekundární zraková korová oblast (area 18, 19) - oblast přilehlá k primární zrakové kůře. Aferentní vlákna přicházejí z primární zrakové kůry a z asociačních jader v pulvinar thalami. Eferentní vlákna jdou zpět do primární zrakové oblasti a do asociačních korových oblastí. Propojení zrakových a asociačních oblastí je nutné ke komplexní analýze zrakových vjemů. Při poškození centra vzniká vizuální agnosie. Postižený vidí, ale nerozpoznává pozorovaný předmět.
Primární sluchová korová oblast (area 41, 42) zahrnuje gyri temporales transversi. Vzruchy přicházející cestou radiatio acustica z corpus geniculatum mediale jsou tonotopicky uspořádány. Eferentní vlákna pokračují do podkorových sluchových center v corpus geniculatum mediale a colliculus inferior. Centrum přijímá sluchové vjemy a jeho poškození vede k postižení sluchu.
Sekundární sluchová korová oblast (area 22) - zevně okolo primární sluchové oblasti. Dostává spoje z primární sluchové kůry asociačních jader v pulvinar thalami. Eferentní vlákna jdou do primární sluchové kůry. Součástí oblasti je Wernickeho sensorické centrum řeči.
Wernickeho sensorické centrum řeči – asociační oblast odpovědná za porozumění mluvené řeči. Při poškození v dominantní hemisféře vzniká stav kdy postižený slyší, ale nerozumí mluvené řeči.
Chuťová korová oblast (area 43) v dolní části pars opercularis gyrus frontalis inferior. Aferenty přicházejí z ncl. gustatorius po přepojení v ncl. ventralis posteromedialis thalami.
Vestibulární korová oblast je rozložena v gyrus postcentralis v oblasti pro hlavu (area 2) a také v gyrus temporalis superior před primární sluchovou kůrou. Dostává impulsy z vestibuárních jader po přepojení v thalamických jádrech. Odstředivá vlákna spojují oblast se zrakovou kůrou a s dalšími oblastmi ovlivňující somato- i visceromotorické systémy.
Primární čichová korová oblast - zahrnuje paleokortex a peripaleokortex v trigonum olfactorium, substantia perforata anterior, část uncus gyri parahippocampalis (area 51), area prepiriformis. Oblast přijímá podněty cestou čichových vláken po přepojení v bulbus olfactorius. Oblast je spojena eferentnímy vlákny s ostatními oblastmi paleokortexu, s limbickou kůrou, s area enthorinalis, amygdalou, hypothalamem. Význam oblasti je v analýze čichových vjemů.
Insulární korová oblast (area 13, 14, 15, 16) – kůra insuly obsahuje přechodné oblasti peripaleocortexu a isocortex. Je považována za centrum gastrointestinální visceromotoriky i viscerosenzitivity.
Limbická korová oblast zahrnuje gyrus paraterminalis, area subcallosa, gyrus cinguli, gyrus parahippocampalis, gyrus dentatus a do postranních komor vyklenutý pes hippocampi, subiculum a struktury archicortexu na corpus callosum. Cytoarchitektonicky jsou tyto oblasti tvořeny archicortexem, periarchicortexem a neocortexem. Aferenní spoje přicházejí z limbické korové oblasti z čichové kůry, z ostatních korových oblastí, z corpus amygdaloideum, septa, thalamu, hypothalamu. Jednotlivé oblasti limbické kůry jsou navzájem dobře propojeny. Eferentní spoje vedou do asociačních korových oblastí, do podkorových částí limbického systému stejné i druhé strany. Funkce limbické korové oblasti vyplývají z jejího zapojení do limbického systému.3
3 Kikalová K., Machálek L., Holibka V., Kutal M., Bezděková M., Limbická korová oblast, NEUROANATOMIE I. - Struktury centrálního nervového systému [online], Univerzita Palackého v Olomouci, Dostupné z WWW: http://www.nan.upol.cz/neuro/cd717.html
Asociační korové oblasti jsou uloženy v jednotlivých lalocích (v oblasti čelního pólu hemisféry, na zevní ploše parietálního laloku, na spánkovém laloku v okolí pólu a na bazální ploše, v týlním laloku v oblasti sekundární zrakové kůry). Asociační oblasti kůry mají reciproční spojení s ostatními korovými oblastmi. Podíl asociačních oblastí na celkové ploše mozkové kůry s evolučním vývojem roste. Zajišťují integraci senzorických podnětů, ovlivňují složité utváření osobnosti – jednání, chování, emoce.
↑ sbalit poznámku ↑
| Obrázky ke kapitole: Koncový mozek – telencephalon | |||

Obr. 47 Laloky koncového mozku
|
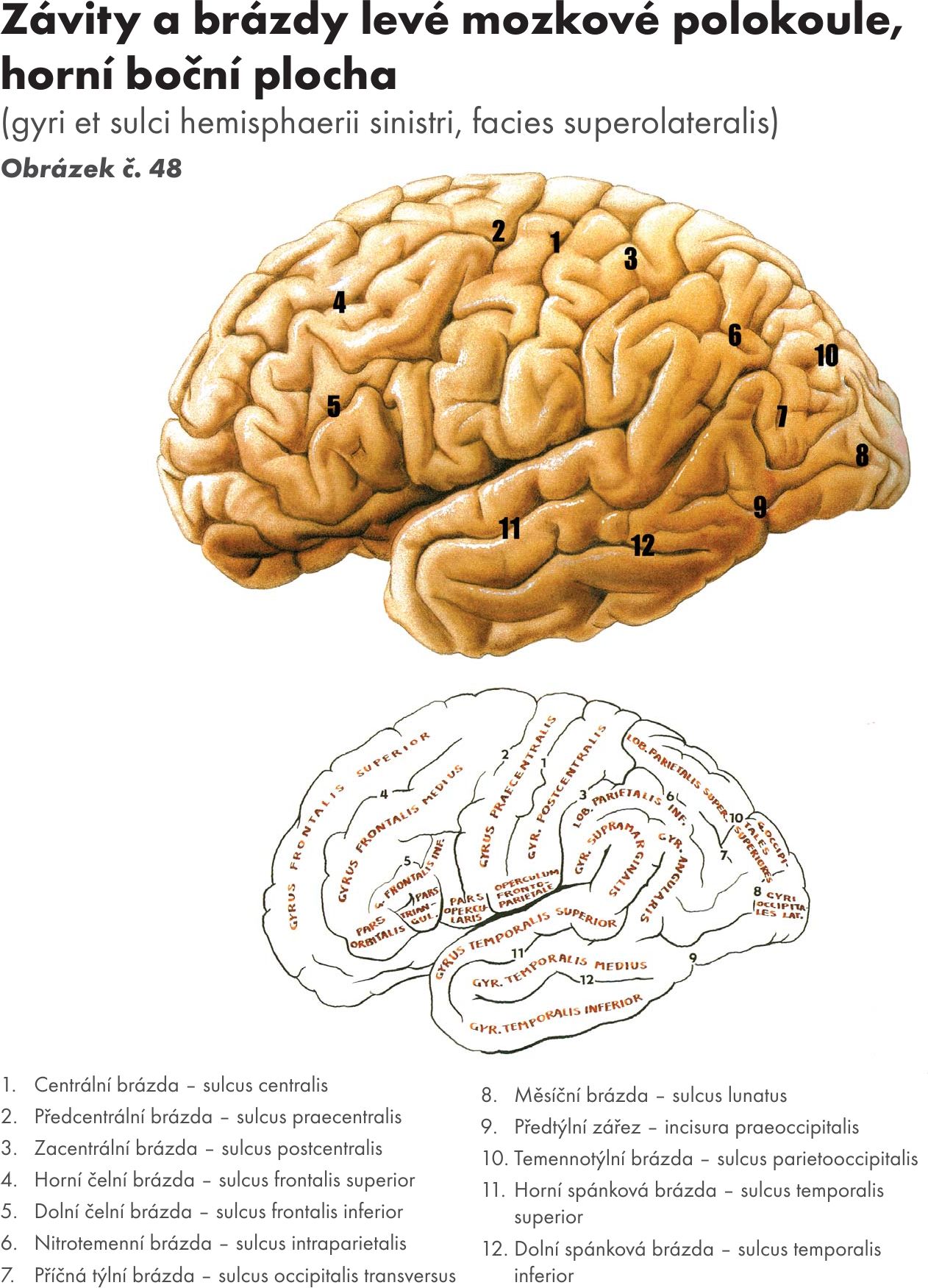
Obr. 48 Závity a brázdy levé mozkové polokoule, horní boční plocha
|
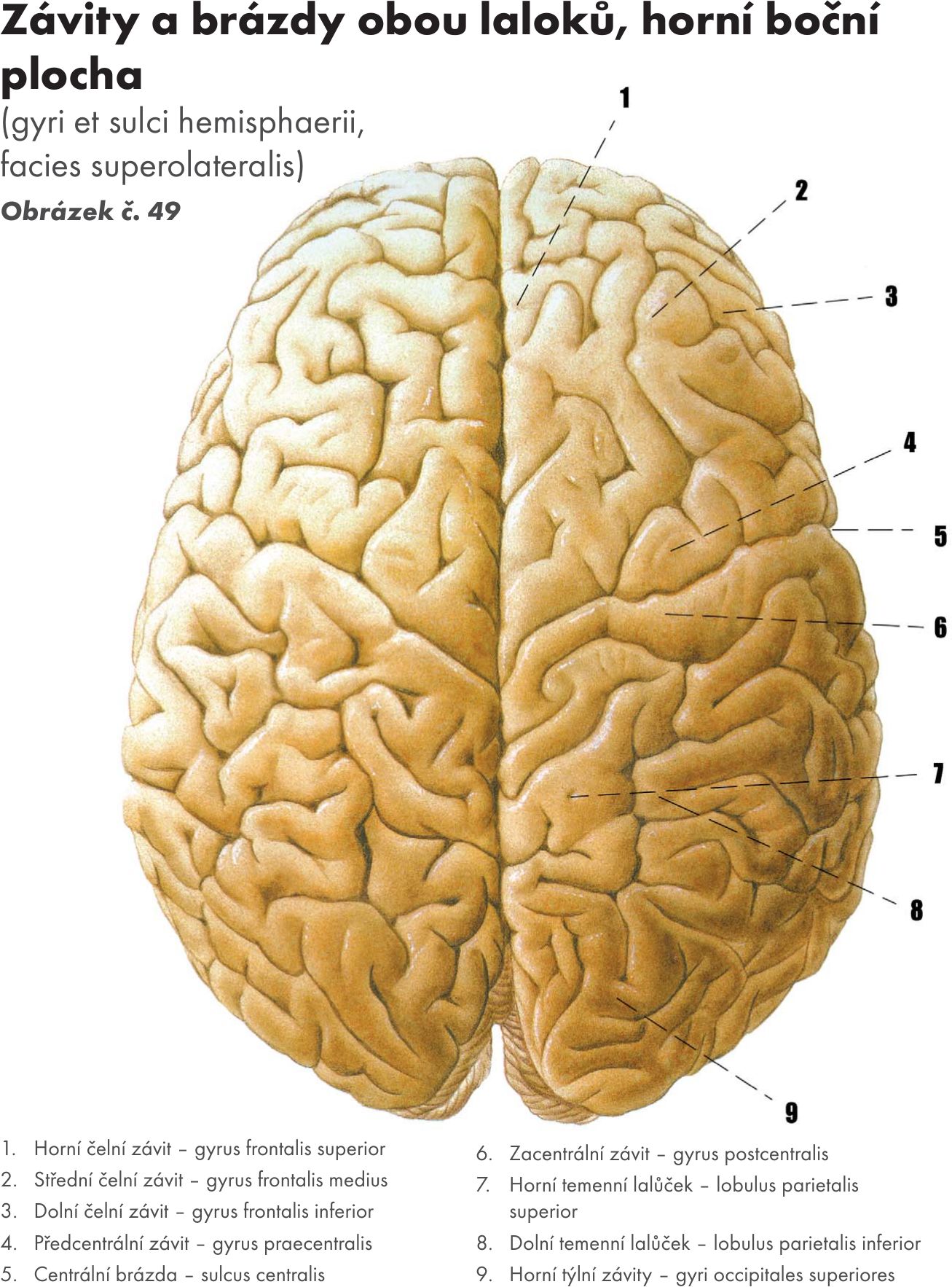
Obr. 49 Závity a brázdy obou laloků, horní boční plocha
|

Obr. 50 Závity a brázdy mozkové polokoule, pravá vnitřní plocha
|
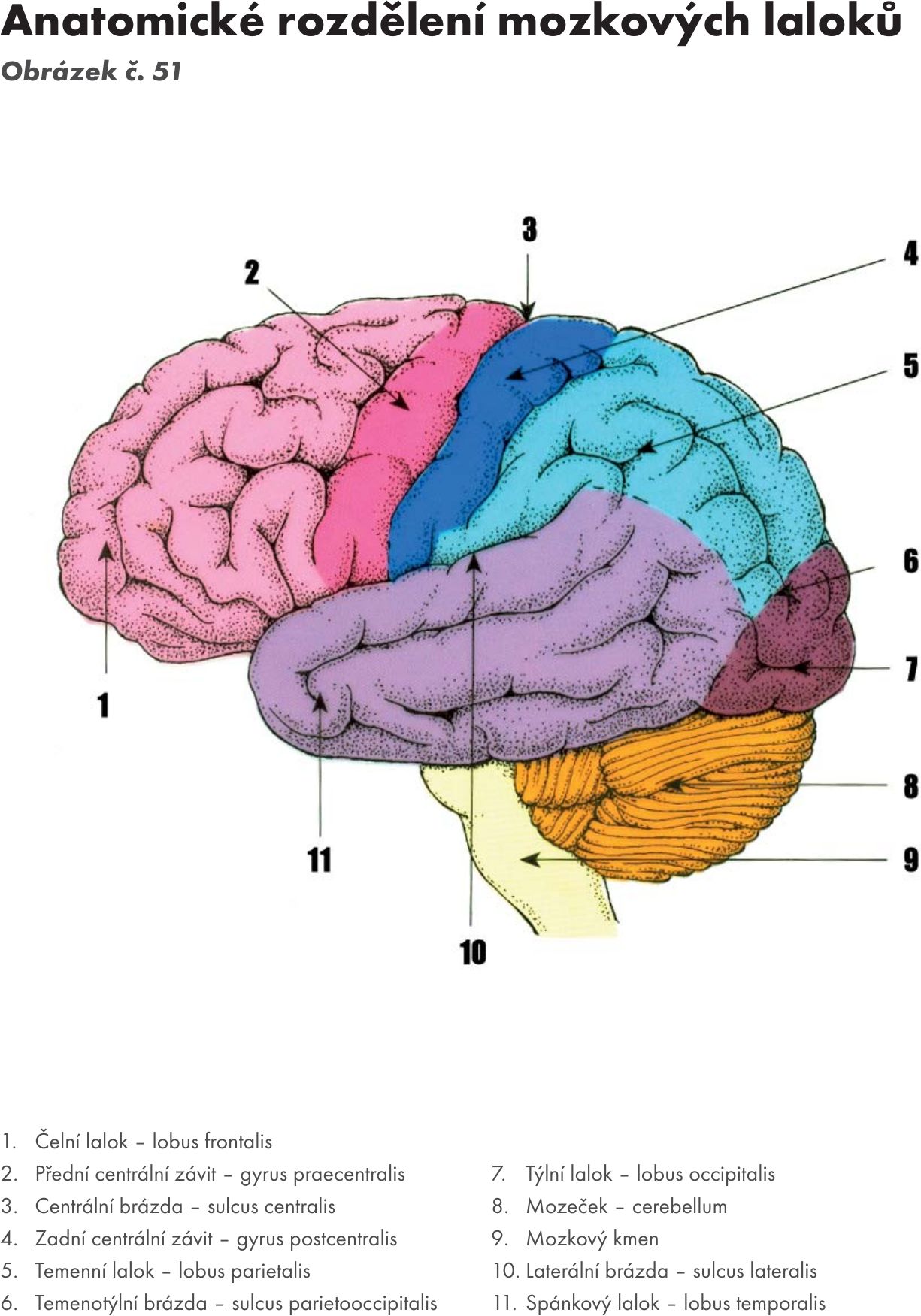
Obr. 51 Anatomické rozdělení mozkových laloků
|
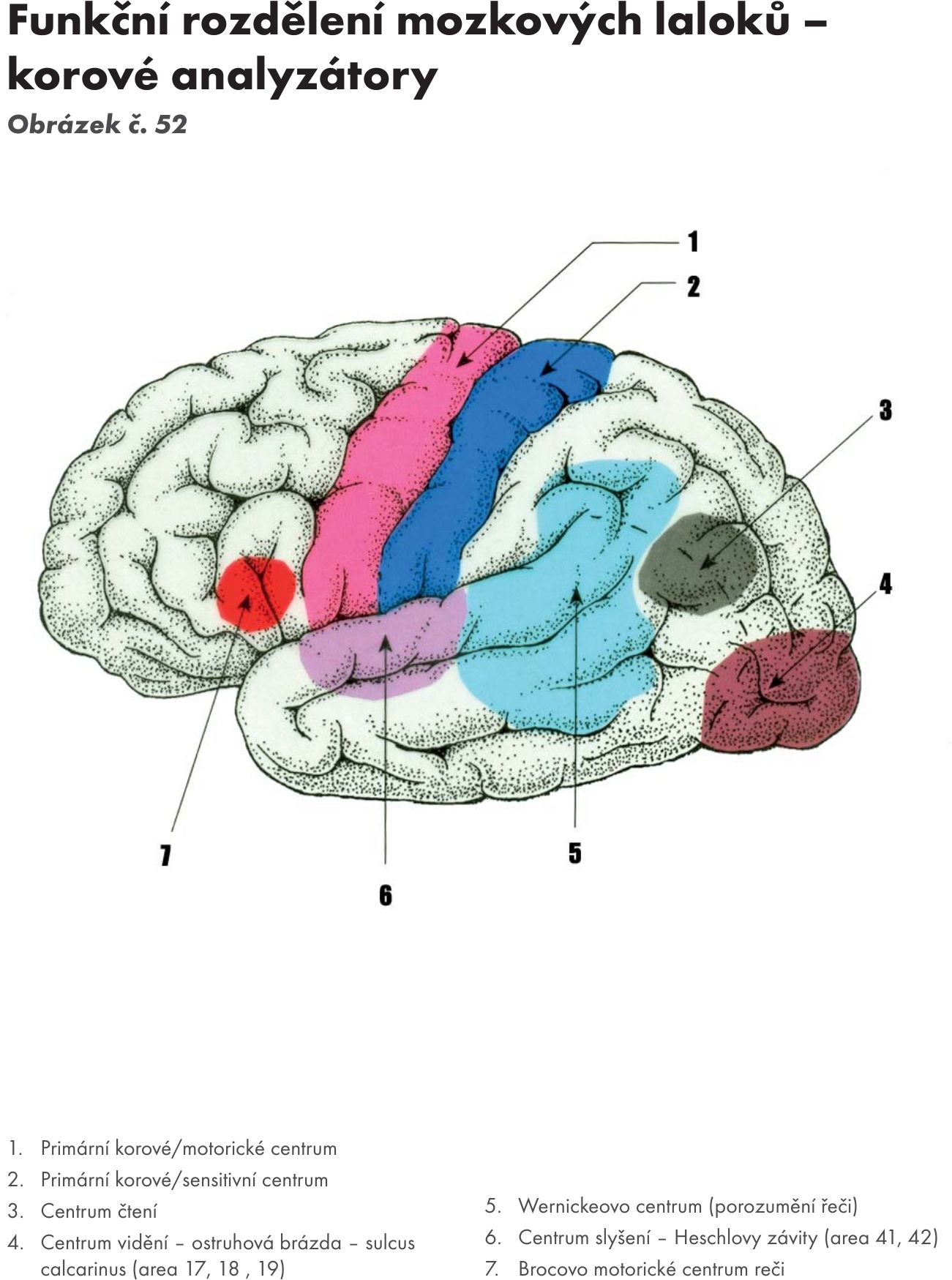
Obr. 52 Funkční rozdělení mozkových laloků – korové analyzátory
|
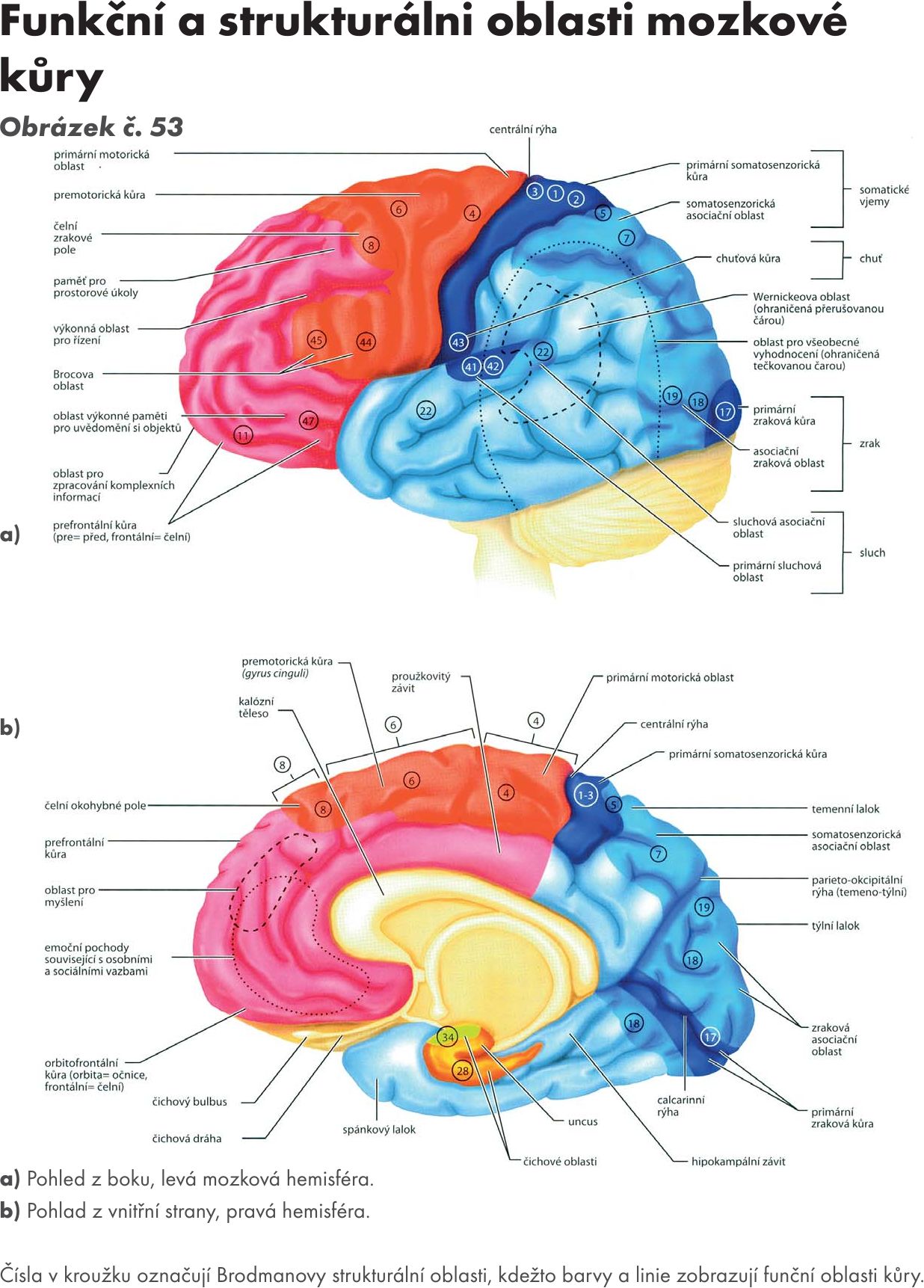
Obr. 53 Funkční a strukturální oblasti mozkové kůry
|
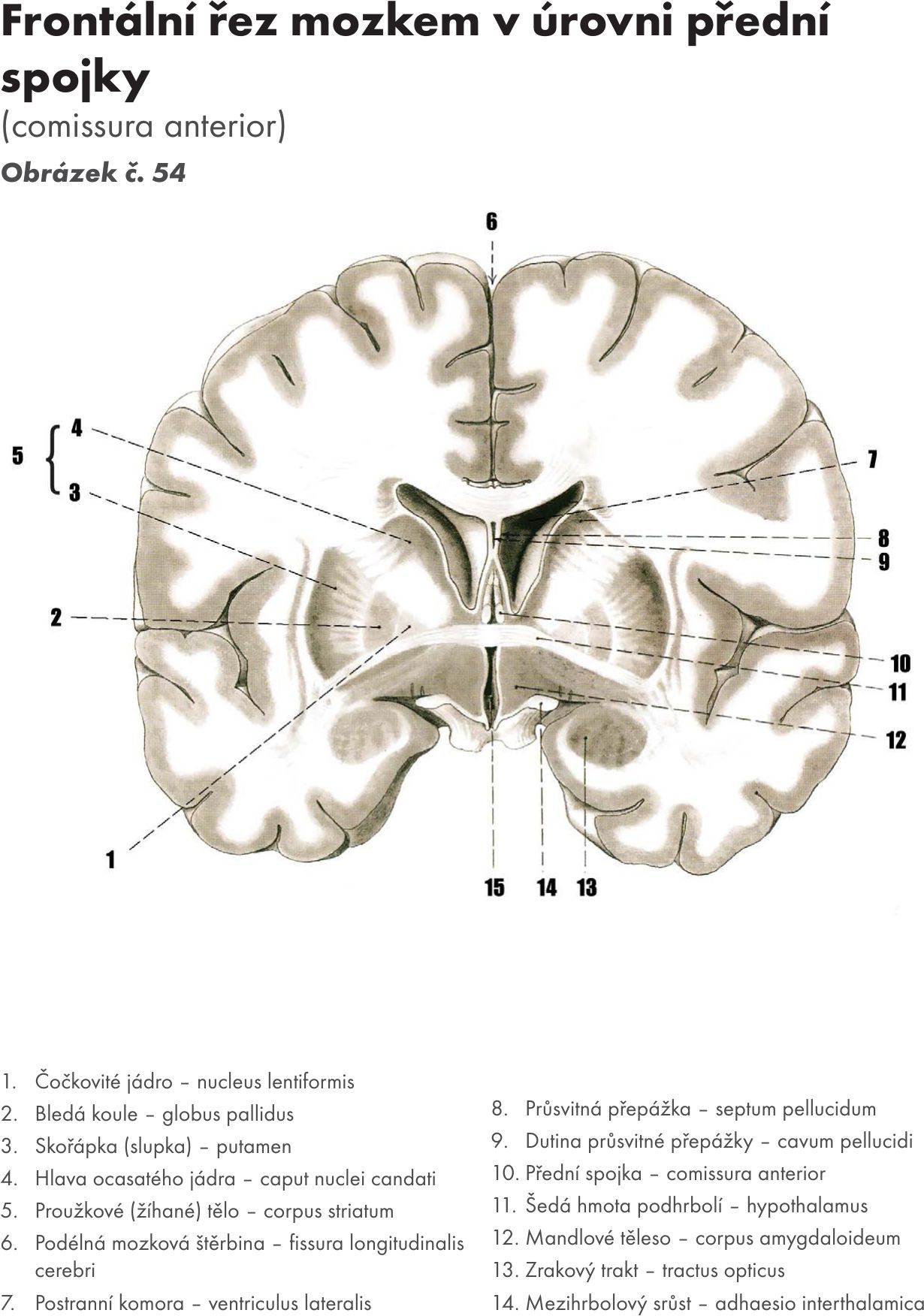
Obr. 54 Frontální řez mozkem v úrovni přední spojky
|
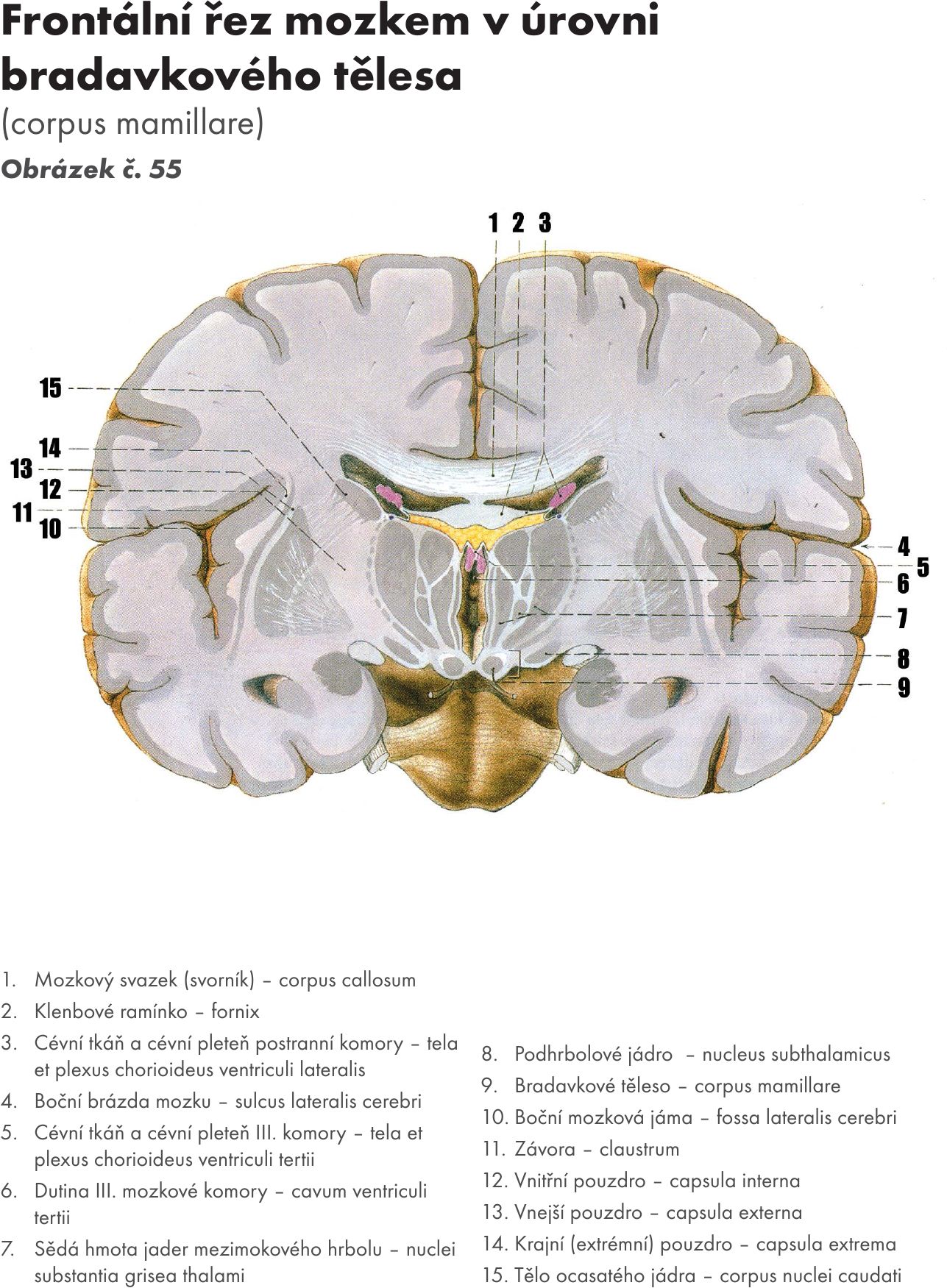
Obr. 55 Frontální řez mozkem v úrovni bradavkového tělesa
|
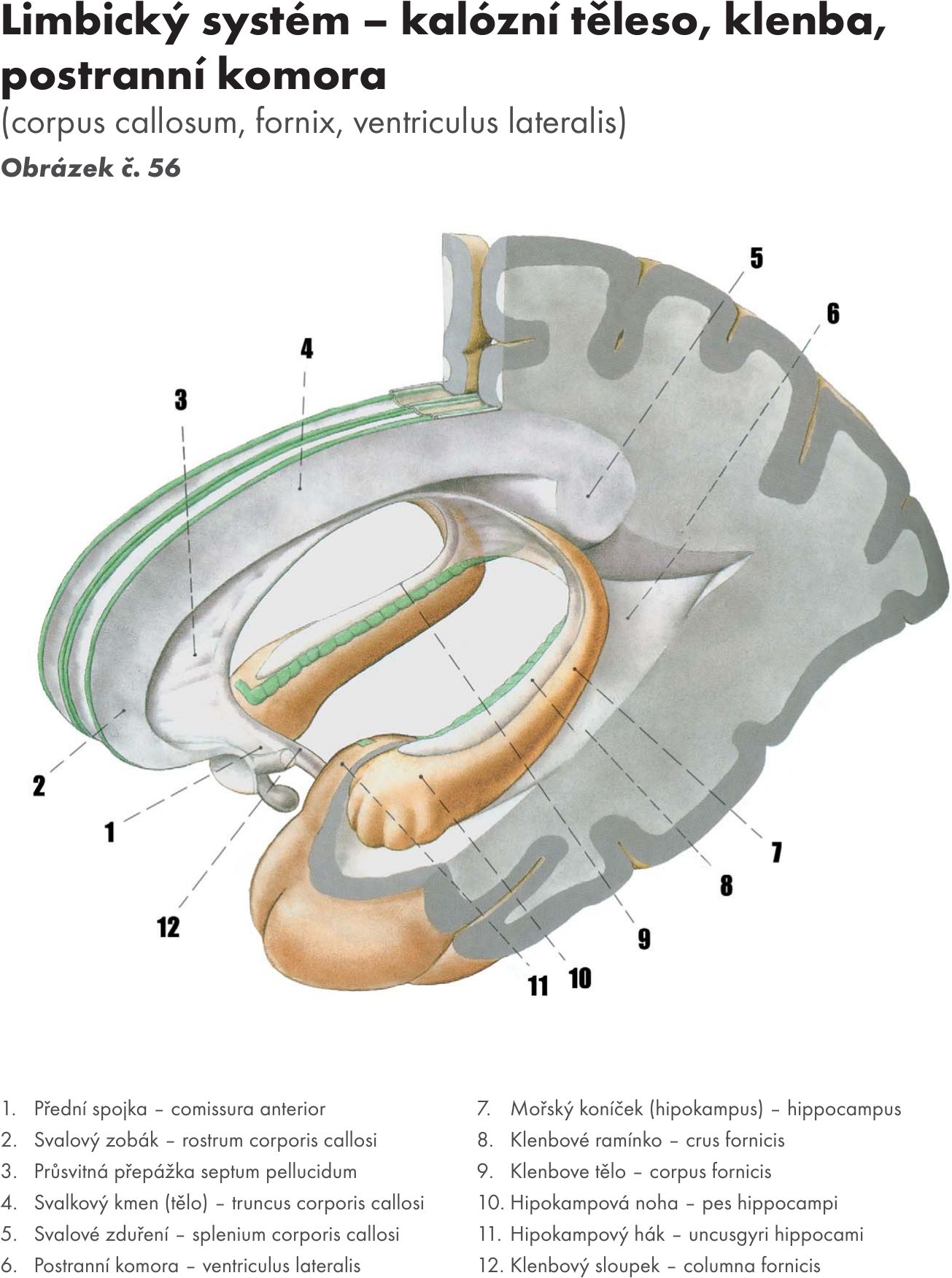
Obr. 56 Limbický systém – kalózní těleso, klenba, postranní komora
|
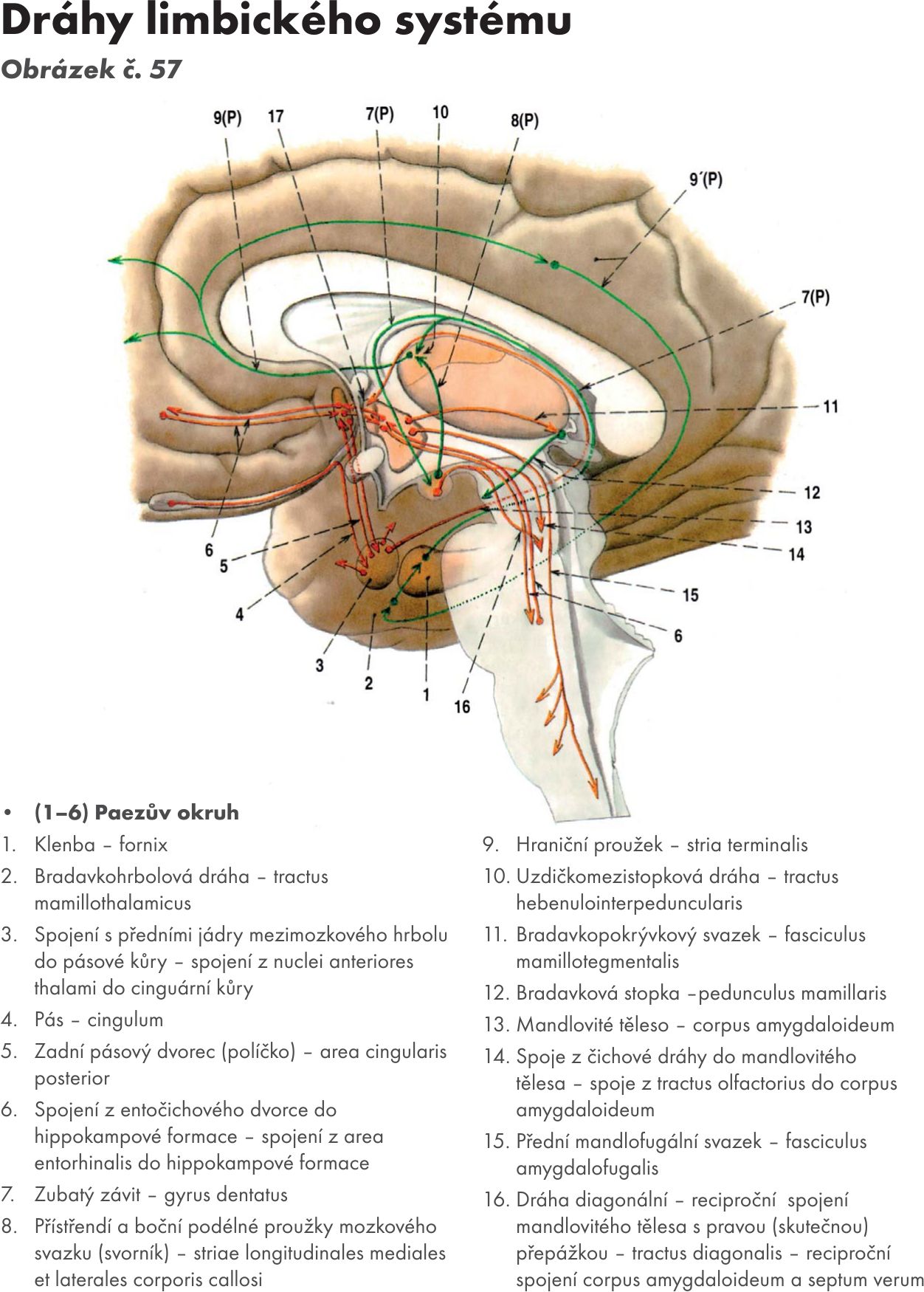
Obr. 57 Dráhy limbického systému
|
|
Dutiny a kanálky centrálního nervstva
Jednoduchý centrální kanálek primitivní nervové trubice se během vývoje v některých místech zúžil nebo naopak změnil v prostorné dutiny – komory.
Komory jsou 4:
- postranní mozkové komory – ventriculi laterales, tvoří pár, jsou v obou hemisférách, spojeny pomocí foramen interventriculare (Monroi) s
- III. mozkovou komorou – ventriculus tertius, která je spojena pomocí mokovodu – aqueductus mesencephali (Sylvii) se
- IV. mozkovou komorou – ventriculus quartus, která komunikuje pomocí 3 otvorů (párovým bočním foramen Luschkae a nepárového foramen Magendi) se subarachnoidální prostorem, ve kterém jsou na bazi vytvořeny basální cisterny, s páteřním kanálem a s centrálním kanálkem míchy – canalis centralis medulae spinalis.
Komory jsou vystlány ependymem a vyplněny aktivním filtrátem krve – mozkomíšním mokem – liquor cerebrospinalis. Ten se tvoří v chorioidálních pleteních – plexus chorioideus. Každý z těchto plexů je tvořen klubíčkem vlásečnic, které jsou obklopeny vrstvou ependymálních buněk. Filrát prostupuje kapilárami ven, prochází přes ependymální buňky – aktivní proces, a dále cirkuluje celým komorových systémem. Odtéká do subarachnoidálního prostoru, kde se přes klky (villi, Pacchionské granulace) pavučnice vstřebává do mízního a částečně do žilního systému.
Postranní mozkové komory – ventriculi laterales. Jsou uloženy uvnitř hemisfér a rozlišujeme na nich čelní rohy – cornua frontale, středovou část – pars centralis, rohy spánkové a týlní – cornua temporale et occipitale. Ve středové části a spánkových rozích je cévní tkáň – tela chorioidea, která obsahuje cévní pleteně – plexus chorioideus. Tvoří ji měkká plena – pia mater a cévní klubíčko, ve kterém se, jak již bylo popsáno, tvoří mozkomíšní mok.
Mezikomorových otvorem – foramen intervetriculare (Monroi) komunikují postranní komory s III. mozkovou komorou.
Třetí mozková komora – ventriculus tertius. Je sagitálně orientovaná štěrbina v oblasti středního mozku. Její laterální ohraničení tvoří mezimozkové hrboly – thalamus, spodinu podhrbolí – hypothalamus. Ve stropu je cévní pleteň – plexus chorioideus ventriculi tertii.
V zadní části se III. komory zužuje do mokovodu – aqueductus mesencephali, canalis Sylvii, který ji vyúsťuje do IV. mozkové komory – ventriculus quartus.
Čtvrtá mozková komora – ventriculus quartus. Je nepárová dutina uložená mezi mozkových kmenem a mozečkem. Její spodinu tvoří kosočtvercová jáma – fossa rhomboidea . Ohraničena je dolní, střední a horní mozečkovou stopkou – pedunculli cerebellares inferiores, medii et superiores. Strop IV. komory – tegmen ventrikuli quartii tvoří přední dřeňová ploténka – velum medullare superius – destičká bíle hmoty rozepjatá mezi horními mozečkovými stopkami – pedunculli cerebellares superiores a dolní dřeňová ploténka- velum medullare inferius, rozepjatá mezi mozečkem a pedunculli cerebellares inferiores. Dřeňové ploténky jsou na komorové straně kryty ependymem, na zevní straně měkkou plenou – pia mater a obsahují cévní tkáň s cévní pletení – tela et plexus chorioideus ventriculi quarti. Komora je vyplněna mozkomíšním mokem, který je produkován z plexus choriroideus (hlavně v postranních komorách).
Ve stropu IV.komory je v distální části otvor – foramen Magendi a v laterálních recesech každé strany komory jsou otvory – foramina Luschkae – kterými mok odtéká do podpavoučnicové dutiny – cavitas subarachnoidales, cisterna magna, do subarachnoidálních cisteren kolem mozkového kmene a na bazi kolem velkého mozku, dále přes velký týlní otvor do páteřního kanálu. Mimo tyto otvory se přímo vstřikuje mok do centrálního kanálku míchy – canalis centralis medullae spinalis, v distální části kosočtvercové jámy.
Mok se vstřebává úměrně své produkci a to převážně na spodině mozku do lymfatických cév a do kavernozního splavu – sinus cavernosus, a v případě zvýšení nitrolebního tlaku dochází ke vstřebávání, jako forma přepadového ventilu, v oblasti klků v tzv Pacchionských granulacích do konvexitárních splavů tvrdé pleny – sinus durae matrix, a to převážně horního šípového splavu – sinus sagitalis superior.
Mok – (liquor) mozek nadlehčuje a chrání před nárazy a otřesy a vytváří kolem mozku hydraulický plášť. Mozek vlastně v mozku plave, což snižuje jeho váhu o 97 % a chrání tak mozek před svou vlastní vahou. Dále má vlastnosti přenosového media, imunologické a vlastnosti termodynamické.
Během dne se z krve, do které se opět vrací, různou cestou, tvoří asi 500 ml mozkomíšního moku. V celém komorovém systému je obsaženo 40–50 ml moku. Celkový objem moku je asi 125–150 ml.
Cirkulace mozkomíšního mozku
Mozkomíšní mok je tvořen v chorioidálních plexech komor (postranních, III. i IV. komory).
Proudění moku začíná v postranních komorách v oblasti spánkových rohů. Mok proudí směrem do čelních rohů a odtud je pohybem mozkové tkáně posunut do foramen Monroi, které se díky interferenčnímu pohybu mozku (interference dechové vlny, pulsní arteriální vlny a jejich pohybech v prostorech mozku), a zároveň díky svému tvaru, prostorové poloze a anatomickému ohraničení (ventrální část tvoří fornix cerebri, zadní pak povrch lůžka – thalamu) chová jako částečně suficientní chlopeň. Mozkomíšní mok je vstřikován do III. komory, která se jednak plněním mokem a jednak pohyby mozku roztahuje do objemu. Pak dochází k jejímu objemovému zmenšování, zejména pohybem lůžek proti sobě, a mok vystřikuje cestou Sylvického aqueduktu do IV. komory. U dětí je akvedukt téměř kruhového průřezu, u dospělých má tvar trojúhelníku se zaoblenými rohy. Při výstřiku moku do IV. komory dochází k jeho navádění pomocí žlábků na její spodině a pomocí jejího prostorového tvaru do foramen Luschkae, jimi do cisteren laterálně kolem kmene, do foramen Magendi, kterým směřuje do velké cisterny, a dále do centrálního kanálku míchy, ve kterém dochází k protiproudnímu pohybu moku. Oblast vstupu do centrálního kanálku míchy se chová jako vývěva na principu Bernouliova jevu. Z hlediska liquorodynamiky je významný velký týlní otvor a cisterna magna ve spolupráci s mozečkovými tonsilami. Vytváří pulsující ventil, chlopeň usměrňující tok liquoru do páteřního kanálu a do mozkovny na spodinu mozku a v poslední části toku po konvexitách mozkových.
Pozn.: Při řadě onemocnění dochází ke změnám ve složení moku. Vyšetření mozkomíšního moku – biochemické, imunologické a cytologické se provádí po odběru moku nabodnutí durálního vaku tvrdé pleny míšní lumbální punkcí nebo vzácněji subokcipitální punkcí, nebo ve speciálních případech přímých odběrem z mozkových komor punkcí komor.
Hydrocephalus (vodnatelnost mozku) je nahromadění mozkomíšního moku v části nebo v celém komorovém systému mozku, které je způsobeno poruchou jeho cirkulace (produkcem, rezorbce, vlastní cirkulace).Hydrocephalus u novorozenců vede ke zvětšení lebky, protože kostní lebeční švy nejsou ještě srostlé. U dospělých tlak moku ničí mozkovou tkáň, protože lebka je již srostlá. Léčba je chirurgická dle typu hydrocephalu (buď se zavádí speciální systém z umělé hmoty – shunt, který odvádí mozkomíšní mok z hlavy extraanatomicky, nebo se provede přemostění místa, kde je zablokována cirkulace – endoskopicky či extranatomickou umělou spojkou).
| Obrázky ke kapitole: Dutiny a kanálky centrálního nervstva | |||
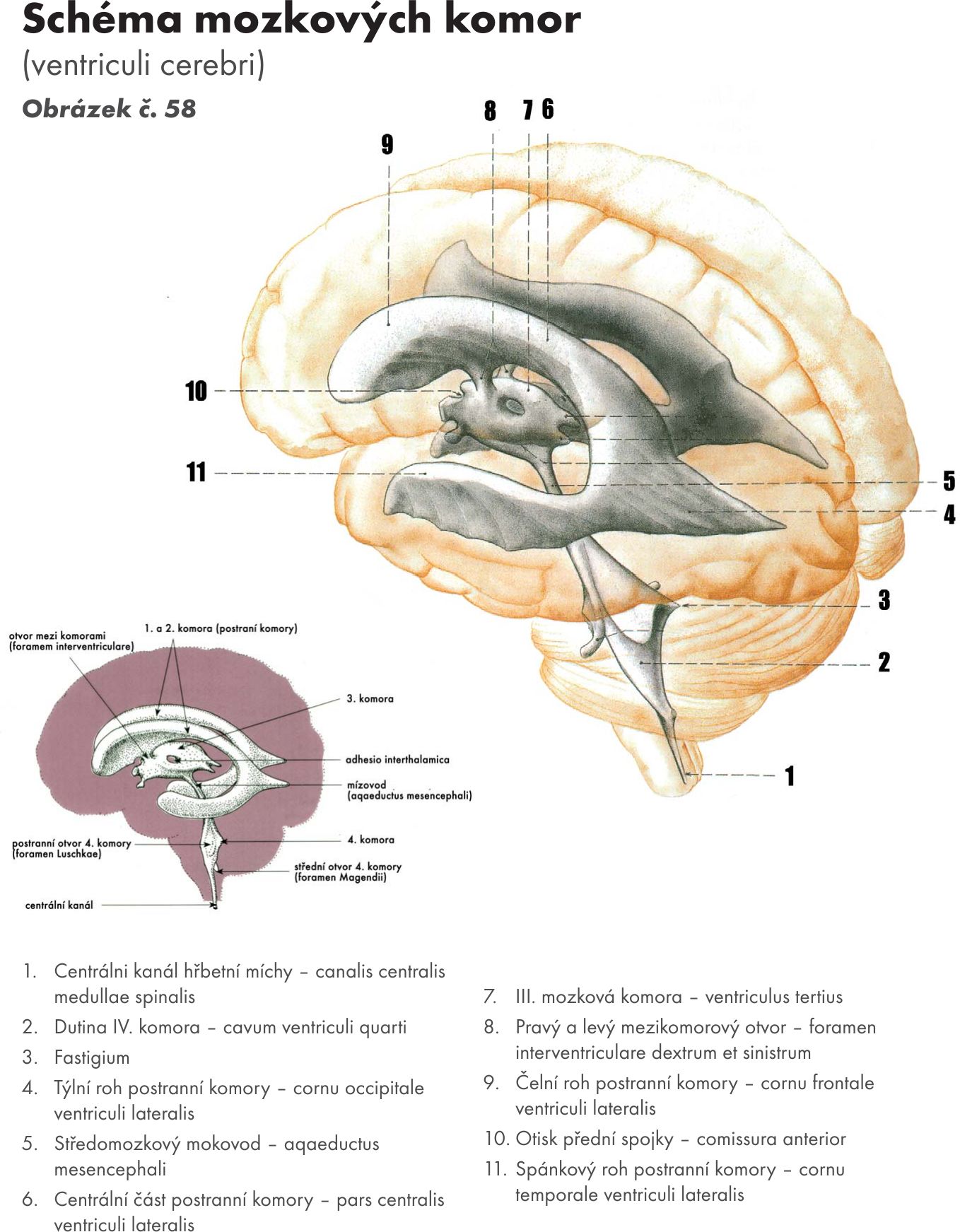
Obr. 58 Schéma mozkových komor
|
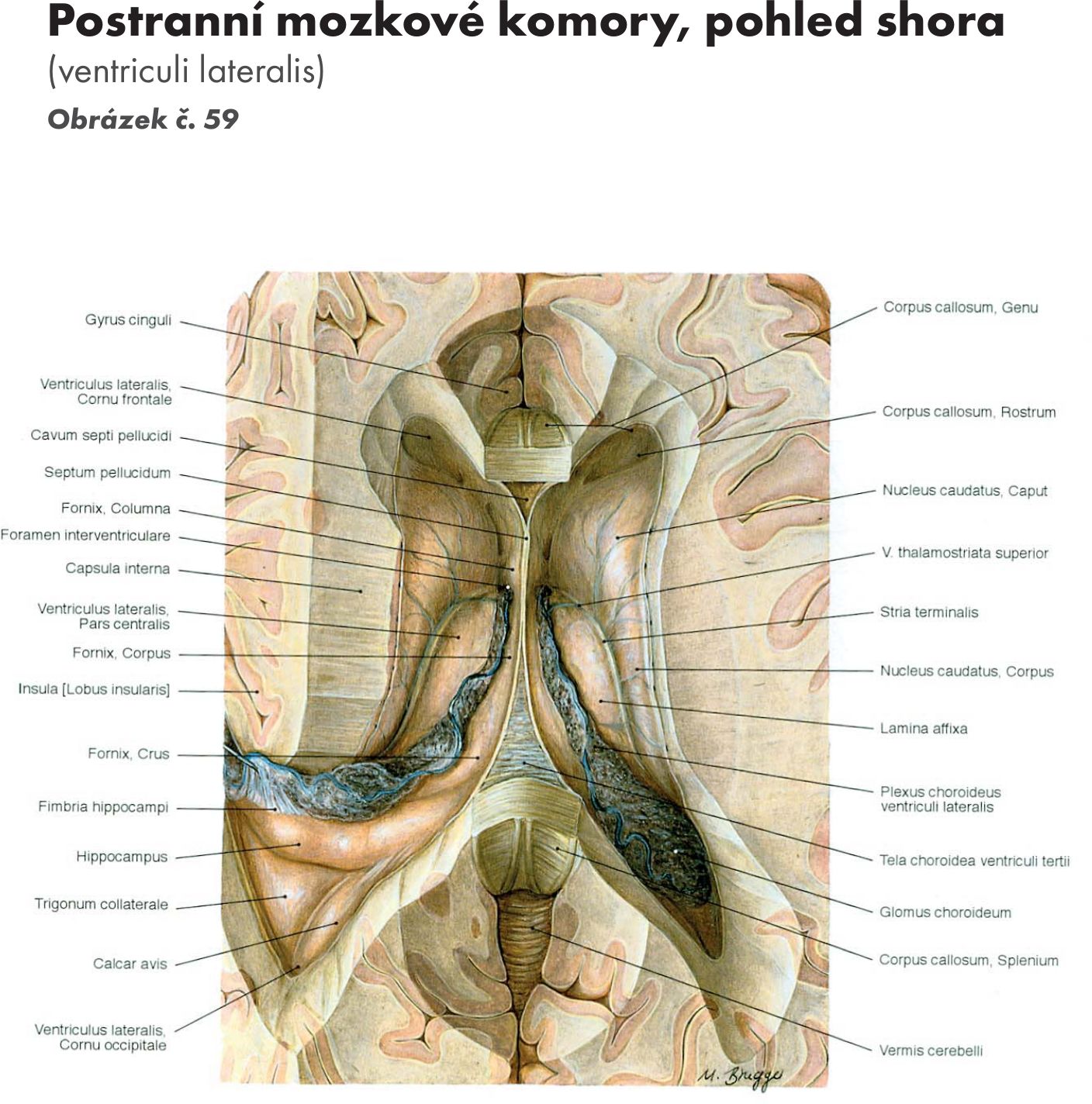
Obr. 59 Postranní mozkové komory, pohled shora
|

Obr. 60 Sekreční buňky (SEM)
|
|
Obaly centrálního nervstva, mozkové pleny – meninges cerebri
Aby nedošlo k poranění měkké a jemné nervové tkáně (poškození nervové buňky jsou již neobnovitelné), je mozek chráněn kostěnou oporou (lebka, páteř) a přílehlými vazivovými obaly – mozkové pleny- meninges a mozkomíšním mokem. Navíce je mozek chráněn proti škodlivým látkám obsaženým v krvi cévně mozkovou bariérou (hematoencephalickou bariérou).
Hematoencefalická bariéra je označení pro bariéru, která odděluje vnitřní prostředí mozku obratlovců od cévního systému v těle a umožňuje jen omezený transport látek mezi mozkovou tkání a krví. Je to možné díky tomu, že buňky cévního endotelu jsou opatřeny výrazně vyvinutými těsnými spoji, jež zamezují vzniku pórů mezi buňkami. Roli hraje do jisté míry i bazální lamina a dále také výběžky astrocytů, které se napojují na vnější stěny krevních vlásečnic a zřejmě vylučují jisté regulační signální proteiny. Přes hematoencefalickou bariéru mohou volně přecházet látky rozpustné v tucích a samozřejmě i plyny (jako je kyslík a oxid uhličitý). Všechny ionty a látky rozpustné ve vodě (jako je glukóza) se však musí aktivně transportovat přes buněčné membrány za spotřeby energie. Díky tomu může být výměna látek mezi krví a mozkovým tkáňovým mokem regulována. Hematoencefalická bariéra v některých oblastech mozku zcela chybí – naopak, v těchto místech se mohou vyskytovat fenestrované kapiláry (s množstvím otvorů v jejich stěnách). Patří k nim epifýza a hypofýza, které vylučují hormony přímo do krve, a některé struktury přiléhající k mozkovým komorám (tzv. cirkumventrikulární struktury, např. area postrema či subfornikální orgán).15
15 Wikipedie: Otevřená encyklopedie: Hematoencefalická bariéra [online]. Dostupný z WWW: http://cs.wikipedia.org/w/index.php?title=Hematoencefalick%C3%A1_bari%C3%A9ra&oldid=10595334
Mozkové pleny jsou tvořeny pojivovou tkání, jsou tři a leží zevně od mozku a míchy a přechází jedna v druhou.
Tvrdá plena – dura mater encephali. Obrázek č. 62 Je nejsilnější z mozkových obalů, tvořená je kolagenním vazivem uspořádáným dle lokalizace. Je tvořena dvěma hlavními vrtvami: buněčnou durální hraniční vrstvou; povrchověji je uložena okosticová vrstva – periostální dura, která srůstá s periostem lebečních kostí, proto není vytvořen epidurální prostor – spatium epidulare. Hlouběji se nachází vlastní tvrdá plena – meningeální dura, která tvoří zevní obal mozku. Tyto dvě vrstvy jsou srostlé mimo místa, kde obkružují žilní dutiny – durální splavy (siny). Mezi tvrdou plenou a lebečními kostmi probíhají plenové artérie a vény (aa. et vv. meningae). Při jejich poranění dochází ke krvácení a postupně odlučování tvrdé pleny od kosti, vytváří se tak sekundárně epidurální prostor a vzniká tzv. epidurální hematom. Je to de facto subperiostální hematom.
Do dutiny lebeční vybíhají řasy – duplikatury tvrdé pleny, které oddělují jednotlivé části – kompartmenty mozku a omezují pohyb mozku uvnitř lebky.
Mozkový srp – falx cerebri. Je široká srpovitá řasa, orientovaná vertikálně a uložená ve střední rovině. Odděluje obě hemisféry mozku. Vpředu se připojuje ke kostnímu hřbětu kosti čichové – crista galli ossis ethmoidalis.
Mozečkový srpek – falx cerebelli. Je také orientován vertikálně a je pokračováním zadní části srpovité řasy mozku směrem dolů. Odděluje obě hemisféry mozečku. Není vždy vyvinut, je velmi částo rudimentální.
Mozečkový stan – (napjatá blána mozečku) – tentorium cerebelli. Je to řasa podobná stanu, je orientována horizontálně a odděluje mozeček od týlních laloků hemisfér.
Sedlová přepážka – diaphragma sellae. Řasa je rozepjatá nad tureckým sedlem – sella turcica. Pod ní v podvěskové jáma – fossa hypophysialis je uložena hypofýza, jejíž stopka prochází otvorem v řase.
Dutina trojklanného nervu – cavitas trigeminale (Meckeli). Leží mezi původním periostem a původní tvrdou plenou. Mezi oběma listy je uložena uzlina n.V.- ganglion trigeminale.
Zevní pochva zrakového nervu – vagina externa nervi optici. Je výchlipkou tvrdé pleny, která obaluje oba zrakové nervy až k oční kouly, kde splývá s bělimou (sclera).
Buněčná durální hraniční vrstva – je tvořena buňkami uspořádanými horizontálně do vrstvev s podélnou štěpitelností těchto vrstev. Na histologickém preparátu se buňky jeví tmavší oproti buňkám dury.
Mozková pavučnice – arachnoidea encephali. Je tenká bezcévná blána uložená pod tvrdou plenou. Přechod mezi oběma je tvořen buněčnou durální vrstvou a arachnoidální buněčnou bariérovou vrstvou. Tato vrtva je tvořena buňkami upořádanými do horizontálních vrstev. Mezi oběma vrstvami je vynikající štěpitelnost a velmi malá soudržnost, což je způsobeno velmi řídkými desmozomálními spoji. Navnitř od této vrstvy přechází arachnoidea do vrstvy arachnoidálních trabekul. Malá soudržnost a pevnost durální hraniční buněčné vrstvy a bariérové arachnoidální vrstvy mezi sebou, umožňuje jejich velmi snadnou separaci, a tím vytvoření tzv. subdurálního prostoru, kde při krvácení pod tvrdou plenu mohou vznikat subdurální hematomy. Vztah mezi těmito vrstvami zároveň umožňuje limitní klouzání mezi sebou.
Tvrdá plena i pavučnice obklopují mozek a nevybíhají do rýh na mozkovém povrchu. Pod pavučnicí je labyrintový prostor – subarachnoidální prostor – spatium subarachnoidale, který je tvořen spletí jemných trámečků arachnoidální tkáně, které připomínají pavoučí sítě. Trámečky neboli arachnoidální trabekuly připevňují pavučnici k pod ní ležící omozečnicí, měkká plena mozková – pia mater. Subarachnoidální prostor je vyplněn mozkomíšním mokem a jsou zde velké krevní cévy zásobující mozek. Rozšířená místa v subarachnoidálním prostoru se nazývají cisterny, jak na konvexitě mozkových hemisféry tak na spodině mozku.
V horní části mozku a na spodině mozku vybíhá pavučnice v arachnoidální výchlipky (klky, villy). Na konvexitě se označují jako Pacchionské granulace – granulationes arachnoidales (Pacchioni). Klky – villy prostupují skrze dura mater se aktivně podílejí na rezorbci mozkomíšního moku ze subarachnoidálního prostoru do lymfatických cév hlavy a do splavů tvrdé pleny mozkové.
Omozečnice – mozková měkká pleny- pia mater encephali. Je vrstva jemné pojivové tkáně, která přesně obemyká a sleduje všechny závity a rýhy mozku. Je bohatě protkána jemnými krevními cévami, které v krátkém průběhu při vstupu do mozkové tkáně rovněž obaluje. V komorách se podílý na vytvoření plexus chorioideus, který produkuje mozkomíšní mok. Krvácení z těchto cév má za následek přítomnost krve v liquoru.
Obaly míchy – meninges spinalis Obrázek č. 63
Podobně jako mozek je i mícha ochráněna kostí – probíhá páteřním kanálem, míšními obaly a mozkomíšním mokem. Míšní obaly jsou jako u mozku tři:
Tvrdá plena míšní – dura mater spinalis se od tvrdé pleny mozku liší tím, že se nepřipojuje k okolním kostem tzn. je bez okostice. Vytváří kolem míchy a měkkých plen míšních vak -saccus durae matris spinalis. Zevně od tvrdé pleny je rozsáhlý epidurální prostor vyplněný tukem a žilními pleteněmi.
Pozn.: do tohoto prostoru se vstřikují anestetika s opioidy pro znecitlivění kořenů míšních a k odstranění pocitů bolesti.
Míšní pavoučnice – arachnoidea spinalis. Je v míšním vaku zevnitř přiložena k tvrdé pleně. Pokračuje přes míšní kořeny na povrch míšních nervů, kde tvoří perineurální epitel.Mezi tvrdou plenou a pavučnicí je velmi intimní kontakt, pavučnice vytváří tří rozsáhlejší kompartmenty kolem míchy, ve kterých se nachází mozkomíšní mok. Nachájecí se zde, podobně jako u pavučnice mozku klky – zejména v oblasti kolem odstupu pochev kořenů – granulationes arachnoidales.
Míšní měkká plena – pia mater spinalis. Je tenká vazivová membrána, která jako u mozku lne k povrchu míchy, kopíruje její povrch a doprovází místní kořeny až do meziobratlových otvorů. Mezi pavučnicí a měkkou plenou je subarachoidnální – podpavoučnicový prostor vyplněný liquorem. Od bočních ploch vysílá zoubkované vazy – ligamenta denticulata, které fixují míchu k tvrdé pleně a vytváří tak její závěsný aparát.
| Obrázky ke kapitole: Obaly centrálního nervstva, mozkové pleny – meninges cerebri | |||
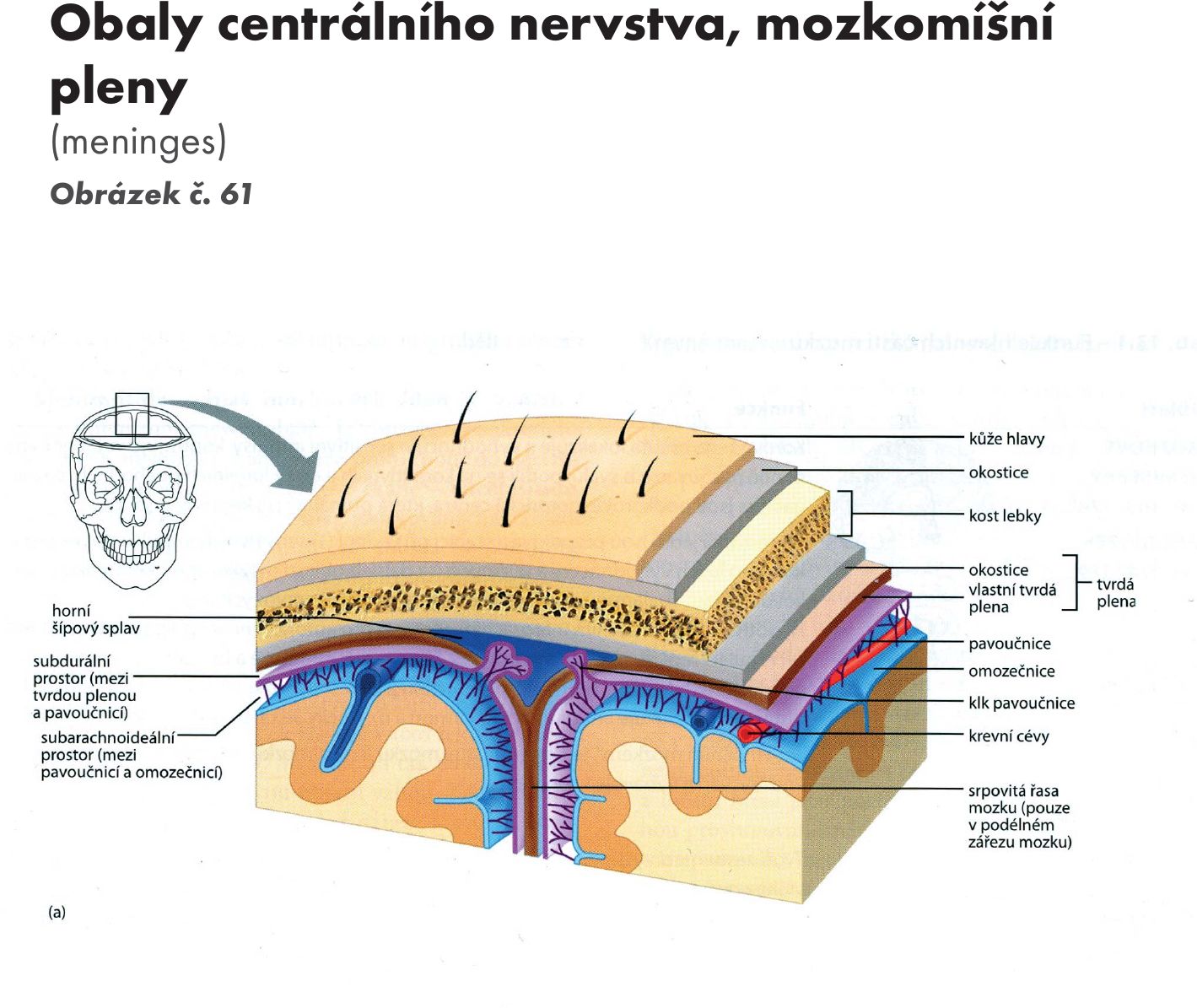
Obr. 61 Obaly centrálního nervstva, mozkomíšní pleny
|
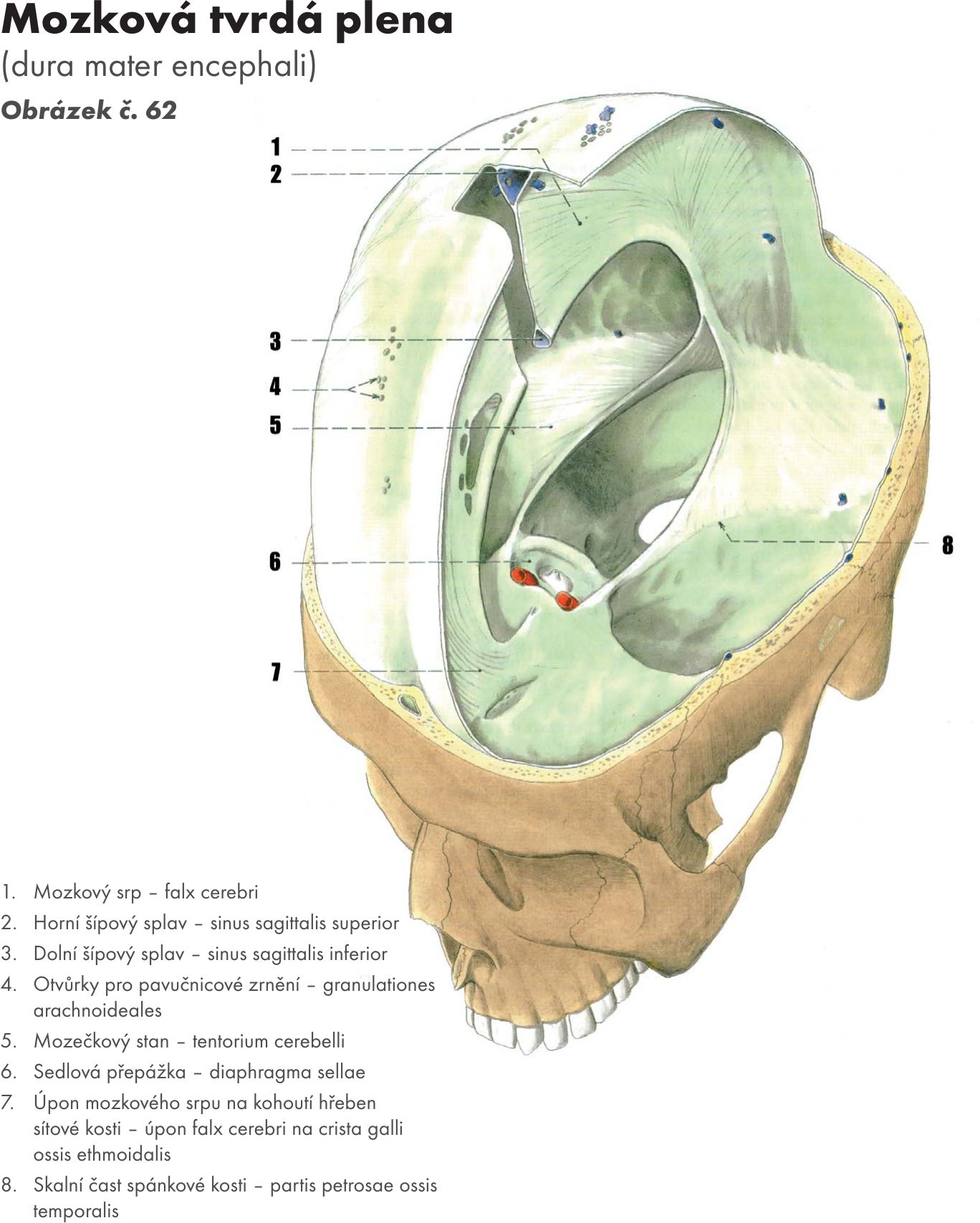
Obr. 62 Mozková tvrdá plena
|
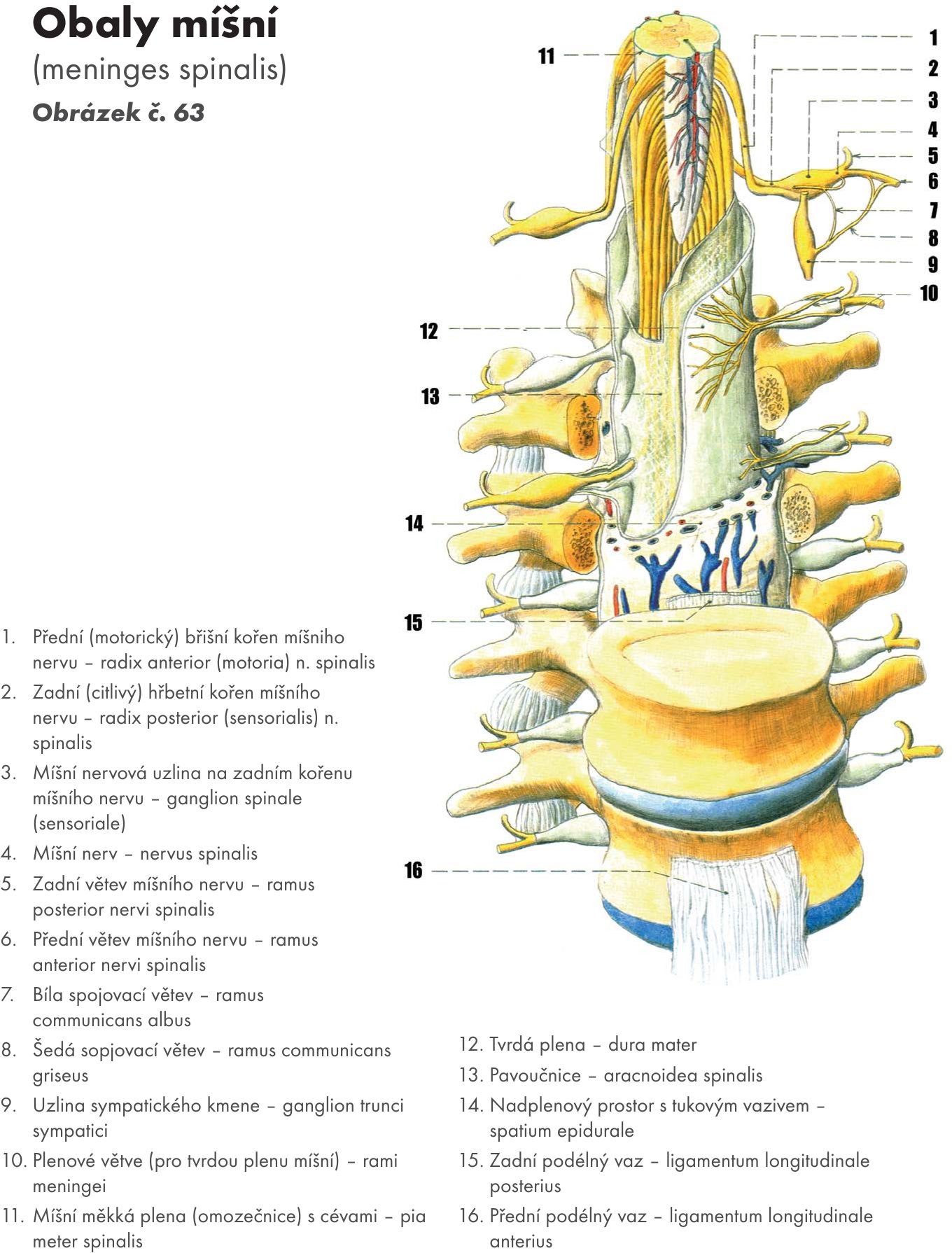
Obr. 63 Obaly míšní
|
|
Cévní zásobení centrálního nervstva (anatomie cévního systému mozku)
Cévní systém mozku je součástí celého kardiovaskulárního systému, ale má svá významná specifika.
Obsahuje část tepennou, žilní a kapilární.
Pozn..:
Historie studia srdce a cévní soustavy je velmi dlouhá. Krom studií v době starého Řecka a Říma, s velkým přerušením v době po zániku Západořímské říše až do renesance. První významnou moderní osobností v jejím studiu byl Vesalius se svým dílem Fabrica: základní popis tělních cév v r. 1543. Pak 1616–1628 Harvey popsal velký a malý oběh a Malpighi popsal kapiláry. 1920 je oceněna práce Krogha Nobelovou cenou za anatomii a fyziologii kapilár. António Caetano de Abreu Freire Egas Moniz r.1927 vyvinul techniku mozkové angiografie, (1949 – nositel Nobelovy ceny za vypracování techniky lobotomie).
Zweifach popisuje roku 1940 mikrocirkulaci a v roce 1967 Rhodin ultrastrukturu mikrocirkulace. Další Nobelovu cenu získává v roce 1980 Palade za studium endotelu a pericytů, které jsou snad kmenové buňky.
Základní oddíly kardiovaskulární soustavy jsou:
- tepny (artérie) – elastického a svalového typu
- tepénky (arterioly) – význam pro distribuci krve a regulaci periferního TK
- vlásečnice (kapiláry) a sinusoidy
- žilky (venuly) a žíly (vény)
- mikrocirkulační řečiště
Celková délka cévní soustavy je cca 40.000 km, cca 200 m2 je plocha endotelu, na který mají vliv endoteliální vasoaktivní faktory.
Důležité je i procento distribuce prokrvení za jednotku času:
- srdce (věnčité tepny) → 5 %
- mozek → 15 %
- svaly → 15 %
- vnitřní orgány → 35 %
- ledviny → 20 %
- kůže, skelet, zbytek těla → 10 %
Průtok mozkem je 15 %, ale váha mozku je 1,3–1,5 kg u průměrného 70 kg člověka, tj. pouze 1,8–2,1 %, objem činí 1350–1700.10-6 m3 a objem člověka 170 cm, 70 kg je 65,9 · 10-3 m3, tj. 2,04–2,6 %.
Cévní systém mozku má významně autonomní pozici v celém kardiovaskulárním systému.
Již vtoková – arteriální část – má svá významná specifika. V přední části je vtok umístěn do dlouhého kostního kanálu, což je pro tepenný systém naprosto netypické, a v zadní části se naopak dvě tepny spojují v jednu, což je rovněž anatomicky jev zcela ojedinělý. Tato výjimečná situace vytváří vstupní část do cévního systému mozku. Ve výtokové části jsou rovněž některá specifika – jednak tenkostěnné systémy prochází různým prostředím z hlediska biomechanických vlastností a jednak jsou dvojího typu: kolabovatelné a nekolabovatelné, které se spolu spojují.
Zcela výjimečnou části cévního systému, anatomickou i fyziologickou, je dutinkový – kavernózní splav – sinus cavernosus, který se označuje v literatuře jako „anatomická šperkovnice“, ale i zároveň jako „šperkovnice fyziologická“. Má několik specifických vlastností:
- tvoří modulátor arteriálního toku – tok se mění do dvou vrstev – zevní proudnice běží po spirále typu α-helixu a vnitřní sloupec proudnic tvoří zúžený sloupec se zrychlením, jako u proudění v tornádu;
- slouží jako pulsní aditivní žilní pumpa, kdy nasává krev z oblasti viscerokrania a vypuzuje krev přes petrózní splavy do jugulární žíly;
- podílí se na resobci likvoru cestou arachnoidálních klků kolem hlavových nervů procházejících splavem (NV,N III.,IV.,V.), přitom se uplatňuje v klcích i tzv. pendel efekt proudění
- slouží jako termoregulátor, či chladič, kdy ochlazenou žilní krví dochází k ochlazení arteriální krve vstupující do neurokrania, čímž se vytváří predispozice pro úpravu termodynamických pochodů mozku. Intrakraniální teplota je oproti teplotě tělesného jádra vyšší za normální teploty cca o 1,5°C. Při rostoucí teplotě tělesného jádra dochází k diferenci až o 3°C. Celý systém kavernózního splavu pak slouží v principu jako vzduchové chlazení, na kterém se podílí paranasální dutiny viscerokrania.
Krom toho se podílí cévní systém v kombinaci s dalšími složkami na „pohybu mozku“ – tedy pulsačních interferenčních pohybech, a to jak povrchu, tak i celého objemu. Jedná se v podstatě o formu prostorové koherence vlnění.
Tepenný systém mozku
Oblast intrakrania je zásobena ze dvou povodí: karotického a vertebrálního, které každé má svá specifika jak anatomická, tak i funkční.
Cévní systém mozku tvoří arteriální část, která běží po povrchu mozku a zanořuje se do mozkové tkáně pomocí perforátorů tepen pronikajících z povrchu do hloubky, a kapilární část, jež se pak sbíhá do žilní části.
Mozek je zásoben z vnitřní krkavice – arteria carotis interna, která je párová, vystupuje z společné krkavice – arteria carotis communis, a dále páteřní tepny – arteria vertebralis, která je rovněž párová.
Vnitřní krkavice – arteria carotis interna se oboustranně dělí na 5 částí. Pod bází leží tepna v měkkých strukturách, pak proniká do kostního kanálku v kosti skalní krkavicového kanálu – canalis caroticus, kterým běží ventromediálně (úsek C5), pak vstupuje do kavernózního splavu (úsek C4), kdy je obejmuta vazivovou manžetou při vstupu – dolní prstenec-ring, poté ostrým ohybem vzhůru opouští kavernózní splav (úsek C3), kdy při výstupu je fixována k okolí pomocí dvou vazivových prstenců – vnitřní a zevní prstenec-ring. Ty se skládají ze dvouvrstvé tvrdé pleny mozkové, která tvoří stěnu kavernózního splavu, po vystoupení z kavernózního splavu vstupuje do subarachnoidálního prostoru (úsek C2) a směřuje okcipitálně, a pak novým ohybem dokončuje úsekem C1 svého esovitého zakřivení nazvané syfon (Moniz).
Pozn.:
Průměr této tepny je 3,3–5,4. 10-3 m, vnitrřní průměr je 3,0–5,5.10-3 m, přičemž vpravo 4,08.10-3 m, vlevo 3,98. 10-3 m.
Pak se vnitřní krkavice dělí na střední tepnu mozkovou – arteria cerebri media a přední tepnu mozkovou – arteria cerebri anterior. Dozadu pod bifurkací odstupuje zadní spojovací tepna – a. communicans posterior a přední tepna komorové pleteně – a. chorioidea anterior, v oblasti v kavernózním splavu odstupuje: oční tepna – a. ophthalmica, boční, zadní a prostřední krkavicodutinkový kmen – truncus caroticocavernosus lateralis, posterior, medialis, přední dolní podvěsková tepna – arteria hypophyseos inferior anterior (Varolli), zadní dolní podvěsková tepna – arteria hypophysealis inferior posterior, horní podvěsková tepna – arteria hypophysealis superior, plenopodvěskový kmen – truncus meningohypophysealis a jeho první větev stanová tepna – arteria tentorii (Bernasconi-Cassinari), střední stanová tepna – arteria tentorii media. Tepny probíhající v kavernózním splavu mají undulující, vlnitý průběh, což umožňuje měnit přímou délku tepny při pohybu vnitřní krkavice – arteria carotis interna v dutinkovém – kavernózním splavu.
Spojením obou předních tepen – arteriae anteriores na spodině mozku ve střední čáře interhemisferálně a obou zadních spojujících tepen – aa. communicantes posteriores spojujích karotické řečiště s řečištěm vertebrálním dotvářejí Willisův tepenný – arteriální okruh (circulus arteriosus Willisi, circulus arteriosus cerebri, heptagon Willisi – 1664). Obrázek č. 64
Korové tepny, které probíhají v měkké pleně – pia mater po povrchu mozku, vyživují mozkovou kůru a přiléhající části bílé hmoty mozkové.
Tepénky basální jsou vesměs velmi drobné, vystupují přímo z tepenného okruhu a zanořují se zespodu do mozku. Živí především basální ganglia s přilehlou bílou hmotou, mezimozek a část středního mozku. Hranice mezi povodím obou tepenných skupin je v bílé hmotě. Artérky obou povodí mají spolu jen nečetné anastomózy.
Tepny komorové pleteně – arteriae chorioideae jsou určeny pro výživu a funkci komorové pleteně – plexus chorioideus komory třetí a komor postranních a přiléhajících částí basálních ganglií. Vyskytují se ve dvou párech.
Přední tepna komorové pleteně – arteria chorioidea anterior vystupuje z vnitřní krkavice – arteria karotis interna v místě pod střední mozkovou tepnou – arteria cerebri media, běží laterálně od zkřížení – chiasmatu po zevní straně zrakového pásma – tractus opticus, do něhož vydává větévky (proto vzniká při jejím uzávěru hemianopsie), klade se přes hák – uncus a vstupuje do cévní tkáně postranní komory – tela chorioidea ventriculi lateralis. Svými větvemi se podílí také na výživě bledého tělesa – globus pallidus a střední části vnitřního pouzdra – capsula interna. V komorové pletení postranní komory – plexus chorioideus ventriculi lateralis se větví v celém jeho rozsahu.
Zadní tepna komorové pleteně – arteria chorirodea posterior začíná z posterolaterální skupiny jako basální artérie. Je velmi často dvojitá. Obkružuje stopku mozkovou -pedunculi cerebri, jde po stranách čtverohrbolí a po straně šišinky – corpus pineale, kterou živí, stejně jako polštář hrbolu – pulvinar thalamu. Větví se ve stropu třetí komory do komorové pleteně třetí komory – plexus chorioideus ventriculi tertii a anastromozuje s větvemi přední tepny komorové pleteně – arteriae chorioidae anteriores.
V přední části z tepen Willisova okruhu vybíhají četné perforující skupiny basálních tepének. Větvičky jsou mnohočetné a vesměs drobné. Lze je rozdělit na 4 skupiny:
Skupina předopřístřední – anteromediální, průměrně 3 cévy (0–10) začínající v oblasti přední mozkové tepny – arteria cerebri anterior a z přední spojovácí tepny – arteria communicant anterior. Cévy se vnořují v přední dírkované hmotě – substantia perforata anterior a zevně od ní. Živí okolí chiasmatu, lamina terminalis, přední část hypothalamu a přední okraj thalamu.
Skupina přednoboční – anterolaterální, průměrně 10 cév (4–21), ze začátku střední mozkové tepny – arteria cerebri media a přední mozkové tepny – arteria cerebri anterior. Vznikají laterálně od přední dírkováné hmoty – substantia perforata anterior a větví se v caput nuclei caudati, v putamen, přední části capsula interna a přední části globus pallidus. Jsou zahrnuty pod název větve žíhané – rami striati, jedna z těchto arterií je značně variabilní, vine se po zevní partií putamen, často dochází k jejímu prasknutí a je označována jako krvácivá tepny – arteria haemorhagica (Charcoti) – tepénka Charcotova. Další vybíhá ze začátku úseku A2 z přední mozkové tepny – arteria cerebri anterior laterálně, jako tepna střední žíhaná – arteria striata media (Strong) a jiná, která živí caput nuclei caudati, se označuje jako tepna hlavní žíhaná – arteria striata princes nebo tepna dlouhá střední – arteria centralis longa či přední návratná tepny – arteria recurrens anterior (Heubneri). Její začátek je velmi variabilní, může vznikat ze skupiny anteroletarální nebo anteromediální. Jednotlivé větvičky dosahují do spodních částí thalamu.
Skupina zadopřístřední – posteromediální, průměrně 4,1 cév (0–15), vychází ze zadní mozkové tepny – arteria cerebri posterior, zčásti též ze začátku zadní spojovací tepny – arteria communicans posterior. Její větve pronikají převážně do zadní dírkované hmoty – substantia perforata posterior. Větví se do thalamu, přední části hypothalamu, ale také do středního mozku, do tegmentum mesencephali a v něm do nucleus ruber.
Skupina zadoboční – posterolaterální, průměrně 8 cév (4–12), začíná v počátku zadní mozkové tepny – arteriae cerebri posterior, zásobuje vedle zadní části thalamu zadní rameno capsula interna a corpora geniculata – hrbolokolenní tepny – arteriae thalamogeniculatae.
Korové tepny jsou v periferním větvení mezi sebou spojeny, ale spojky se zpravidla neuplatní při uzávěru tepny v začátku hlavních korových tepen.
Hlavní korové tepny mají 3 páry:
Přední mozková tepna – arteria cerebri anterior. Obrázek č. 65 Přední mozková tepna – arteria cerebri anterior běží z vnitřní karotidy v intrakraniu subarachoidálními cisternami na bázi mozku mediálním směrem, téměř paralelně se spodinou lebeční, Na spodině mozku ke střední čáře, označuje se jako úsek A1. V průběhu vydává četné perforující tepny do hlubokých struktur mozku a dále větve na spodinu čelního laloku. Přední mozková tepna – arteria cerebri anterior se mediánně spojuje s druhostrannou přes přední spojovací tepnu – a. communicans anterior, která dále běží mezi oběma hemisférami ventrorostálně, označuje se jako úsek A2, poté obloukem kolem corpus callosum, úsek A3, se stáčí pak kraniokaudálně směrem okcipitálním, kde se nazývá okolosvalková tepna – arteria pericallosa. Z nich pak vybíhají četné cévy na mediální plochu mozkových hemisfér, např. arteria cingulomarginalis. V každém dalším větvení se úsek označuje jako A4, A5 atd.. Svými větvemi zásobuje oblasti v kůře mediální část orbitální plochy čelního laloku, přes hranu hemisféry zasahuje na zevní plochu a zásobuje gyrus frontalis superior a variabilní část gyrus frontalis medius. Dále zásobuje horní čtvrtinu gyrus praecentralis i postcentralis (tím korové okrsky pro dolní končetinu), horní partie lobulus parietalis superior a celou mediální plochu hemisféry. Přední mozková tepna – arteria cerebri anterior končí v oblasti předklíní – praecuneus, kde se stýká s oblastí zásobení ze zadní mozkové tepny – arteria cerebri posterior.
Střední mozková tepny – arteria cerebri media Obrázek č. 66 je přímým pokračováním vnitřní krkavice – arteria carotis interna. Vkládá se do boční mozkové jámy – fossa lateralis cerebri (Sylvii) a zde se dělí na větší počet větví, které se rozbíhají po čelním, temenním, spánkovém, i na přední okraj laloku týlního. Tak zásobují větší část gyri orbitales, gyrus frontalis inferior, variabilní část gyrus frontalis medius, dolní tři čtvrtiny gyrus praecentralis i postcentralis, gyrus supramarginalis, gyrus angularis, větší část temenního laloku – gyrus temporalis superior, medius a částečně i inferior – a pól temporálního laloku. Z praktického hlediska popisu se střední mozková tepna – arteria cerebri media rovněž člení na úseky – první po dostupu z arteria carotis interna je M1, prvním větvením se mění na M2 a dále M3, M4 atd. V oblasti větvení mezi úseky M2–3 vzniká perforující tepna na mediální ploše pronikající do oblasti capsula interna, externa a extrema, claustra a striata.
Zadní mozková tepny – arteria cerebri posterior, která tvoří zadní část Willisova okruhu, vzniká z povodí basilární tepny, tedy vertebrálního povodí. Tvoří poslední větve bazální tepny – arteria basilaris. Zatáčí vzad okolo pedunculus cerebri, vydává větve do čtverohrbolí a větví se na spodní ploše laloku spánkového a téměř po celém týlním laloku. Samostatnější větví je ostruhová tepna – arteria calcarina, která se dělí v primárním korovém zrakovém centru. Průběh tepny je z praktického hlediska také členěn na úseky – první po větvení z bazální tepny – arteria basilaris je úsek P1, další větvení je úsek P2, 3, 4 atd. Obrázek č. 67
Páteřní tepny – arteriae vertebrales začínají po odstupu z podkličkové tepny – arteria subclavia jako jejich první větve, a pak vstupují v oblasti C6, výjimečně C5, do kostěných žebropříčných otvorů – foramina costotransversaria krčních obratlů, takže jsou uloženy v lůžku za sebou do série řazených 4–5 kostních prstenců objímajících obě tepny, v jejich okolí se nalézá žilní pleteň, která obě tepny doprovází i při průchodu kostními prstenci (pulsující pružná trubice obalená pletení tenkostěnných trubic v tenké vrstvě a na ni navlečených pět kostěných prstenců – tlumič?). Pak vytváří tepny oblouk dorsálně v oblasti C3–2 a v oblasti průchodu otvorem atlasu, ohýbají se medioventrálně a leží ve žlábku vytvořeném na laterální části oblouku atlasu (C1) a pak pronikají několikavrstevnou vazivovou membránou atlantookcipitální v tvrdé pleně, kterou prochází. Tvrdá plena zde tvoří vazivový prstenec kolem tepny. Po proniknutí intrakraniálně do subarachonidálního prostoru a cisteren této oblasti se na přední ploše mozkového kmene spojují v jedinou tepnu arterii basilární – je to výjimečný typ spojení dvou tepen do jediné, běžně se tepny větví.
Pozn.:
Průměr artérie basilární je 3,0 (2,5–4,3) 10-3 m, přičemž zevní 3,81 (2,16–5,52)·10-3 m, vnitřní průměr 3,2(1,73–4,85)·10-3 m, u artérie vertebrální je průměr vnitřní 4,5·10-3 m, vpravo 4,3·10-3 m, vlevo 4,7·10-3 m.
Tepna basální vybíhá po přední ploše mozkového kmene kraniálně a z ní vycházejí tepny – zadní mozečková tepna – a. cerebelli posterior oboustranně, tepny prodloužení míchy- arteriae medullae oblongatae, mostu – arteriae pontis, přední mozečková tepny – arteria cerebrelli anterior oboustranně a horní mozečkové tepny – arteriae cerebelli superiores oboustranně, a následně pak četných perforátorů (25–40) do fossa intercruralis. Basilární tepna se větví na dvě své konečné zadní mozkové tepny – arteriae cerebri posteriores a spojuje se tak s tepnami z povodí karotického do Willisova arteriálního okruhu na spodině lebeční. Vlastní výstup všech velkých tepen vstupujících do okruhu – arteriae carotis internae oboustranně a arteria basilaris tvoří trojúhelník malé plochy na spodině mozku. Jedná se o rovnostranný trojúhelník o straně 15–20·10-3 m.
Od tohoto trojúhelníku je délka směrem frontálně, rostrálně a směrem okcipitálně kaudálně stejná – 80–85·10-3 m.
Žilní systém mozku
Dělení žilního systému
Mozkové žíly se dělí na dva typy:
tenkostěnné – kolabovatelné. Ve vztahu k prostředí, v němž se tenkostěnné kolabovatelné trubice nacházejí, jej lze rozčlenit do 3 skupin: intracerebrální, subpiální či subependymové a přemosťující žíly.
silnostěnné, tuhé – splavy mozkové, jejichž stěna je tvořena tvrdou plenou mozkovou, jsou velmi málo poddajné, nekolabovatelné.
Vzhledem k tomu, že žíly vybíhají na povrch mozkové tkáně nejen na konvexitě, ale i na spodině mozku a v komorách, dělí se žilní systém mozku z anatomického pohledu na: povrchový a hluboký.
Pozn.:
Na základě studie histologické lze rozdělit výtokové části přemosťujících žil na 4 typy.
Dělení výtokové části přemosťujících žil:
- krytí arachnoidálními klky,
- se zvětšením průřezu,
- se zmenšením průřezu
- s vytvořenou chlopní (chorda Willisi).
K povrchovým žilám se řadí:
- Horní mozkové žíly – venae cerebri superiores, které po zevní ploše hemisféry sbírají krev, jdou šikmo vzhůru a ústí do horního šípového splavu – sinus sagitalis superior.
- Povrchová střední mozková žíla – vena cerebri media superficialis, je uložena v oblasti Sylvické brázdy – boční mozkové jámy – fossa cerebri lateralis. Ústí do předního či zadního okraje dutinkového splavu – sinus cavernosus nebo do nekonstatního klínotemenního splavu – sinus sphenoparietalis (Breschetii). Sbírá krev ze zevní strany hemisféry. Velmi často tvoří anastomosu s horní mozkovou žílou – venoa cerebri superior kolem precentrální či postcentrální krajiny, čímž vzniká horní, velká spojovací žíla – vena anastomotica superior, magna (Trolard) – Trolardova žíla. Je to přímá spojka mezi dutinkovým splavem – sinus cavernosus a horním šípovým splavem – sinus sagitalis superior. Velmi často je vyvinutá další žilní spojka mezi povrchní střední mozkovou žílou – vena cerebri media superficialis a dolní povrchní spánkovou žílou – vena temporalis superficialis inferior (Labbé) – Labbého žíla, spojující dutinkový splav – sinus cavernosus se splavem příčným – sinus transversus. Povrchová spánková žíla – vena temporalis superficialis ústí velmi variabilně do příčného splavu – sinus transversus, esovitého splavu – sinus sigmoideus či do stanového splavu – tentoriálního sinu. Je velmi často vícečetná a v zadní části mnohdy přechází do dolní povrchové týlní žíly – vena occipitalis superficialis inferior.
- Hluboká střední mozková žíla – vena cerebri media profunda leží hluboku ve boční mozkové jámě – fossa cerebri lateralis a běží paralelně s tepnou. Přijímá přítoky z krajiny operkulární i insulární, má spojky s povrchovou mozkovou střední žílou – vena cerebri media superficialis a ústí do bazální žíly – vena basalis (Rosenthali).
- Dolní mozkové žíly – venae cerebri inferiores sbírají krev na bázi mozkové a vyúsťují do horního skalního splavu – sinus petrosus superior a příčného splavu – sinus transverusus (dolní povrchní spánková žíla – vena temporalis superficialis inferior, dolní povrchní týlní žíla – vena occipitalis superficialis inferior atd.), část žil sbírá krev na spodní ploše čelního laloku v krajině přední dirkované hmoty – substantia perforata anterior a postupně konstituuje žílu, která obemyká penduculi cerebri, přibírá větve ze spodní plochy hypothalamu i z fossa interpeducularis. Tato žíla se nazývá bazální žíla – vena basalis (Rosenthali) a ústí do začátku velké mozkové žíly -vena magna cerebri (Galleni). Vena Rosenthali či vena basalis se tvoří z hluboké cerebrální žíly a přední cerebrální žíly laterálně od vnitřní krkavice při horním okraji dutinkový splav – sinus cavernosus, častěji je spojení ventrální vlevo než vpravo. Žíly – hluboká, cerebrální, mediální a přední, drénují insulární oblast, hypothalamickou přední oblast, laminu terminalis, gyrus rectus a okolní struktury. Spojují se v oblasti rostrální části substantia perforata. Do hluboké cerebrální žíly ústí dolní hrboložíhané žíly – venae thalamostriatae inferiores, především dolní centrální větve, které drénují capsula externa, přední část putamen, capsula interna, hlavu nucleus caudatus a laterální hypothalamickou oblast. Subependymálními žilami v laterálním komoře mozkové se tvoří intracerebrální anastomosy spolu s horní hrboložíhaná žíla – vena thalamostriata superior, to tvoří žilní komunikaci mezi povrchovým a hlubokým systémem.
- Bazální žíla – vena basalis Obrázek č. 68 spolu s venae cerebri anteriores a dalšími větvemi tvoří v oblasti spodiny mozku laterálně od tepen další cévní okruh venozní okruh Trolardův – circulus venosus Trolardi. Tento je žilní paralelou tepenného kruhu Willisova – circulus arteriosus Willisi.
Dále se v rámci povrchového žilního systému vyskytují 2 významné anastomosy:
Trolardova anastomosa (horní, velká spojovací žíla – vena anastomotica superior, magna) – přímá spojka mezi dutinkovým splavem – sinus cavernosus a horním šípovým splavem – sinus sagitalis superior.
Labbého anastomosa (dolní spojovací žíla – vena anastomotica inferior Labbé) – spojka dutinkovým splavem – sinus cavernosus s příčným splavem – sinus transversus.
Hluboké žíly mozkové Obrázek č. 69
Hluboké mozkové žíly – venae cerebri internae jsou dvě žíly uložené na stropině III. mozkové komory. Každá z nich je tvořena soutokem tří žil:
- přední a zadní žíla průsvitné přepážky – vena septi pellucidi, anterior a posterior, která sbírá krev z průsvitné přepážky – septum pellucidum a z hlavy ocasatého jádra – caput nuclei caudati,
- horní žíla komorové pleteně – vena chorioidea superior, která sbírá krev z komorové pleteně postranní komory – plexus chorioideus ventriculi lateralis,
- horní hrboložíhaná žíla – vena thalamostriata, superior (hraniční žíla - vena terminalis), prosvítající v hraničním proužku – stria terminalis a sbírající krev z corpus striatum, z capsula interna a největší části thalamu. Do ní se sbírají přístřední předsíňová žíla postranní komory – vena atrii ventriculi lateralis medialis a boční předsíňová žíla postranní komory – vena atrii lateralis ventriculi lateralis, žíly ocasatého jádra – venae nuclei caudati.
Obě vnitřní žíly – venae internae se spojují pod splenium corporis callosi v nepárovou velkou mozkovou žílu – vena magna cerebri (Galleni), která ústí do přímého splavu – sinus rectus. Na spodní straně velké mozkové žíly – vena cerebri magna je vaskularizované ztluštění arachnoidey, tzv. nadšišinkové pavoučnicové tělo – corpus arachnoideae suprapineale (Clark Le Gros). Do velké mozkové žíly – vena cerebri magna ještě ústí boční přímé žíly – venae directae laterales, zadní žíla mozkového svalku – vena corporis callosi posterior a hřbetní žíla mozkového svalku – vena corporis callosi dorsalis.
Žíly mozečkové lze rozdělit do dvou skupin: Obrázek č. 70
horní mozečkové žíly – venae cerebelli superiores, horní žíly polokoule – venae hemispheriae superiores, hroní červová žíla – vena vermis superior, na horní ploše mozečku, které odvádějí krev mediálně do velké mozkové žily – vena cerebri magna, laterálně do příčného splavu – sinus transversus a horního skalního splavu – sinus petrosus superior,
dolní mozečkové žíly – venae cerebelli inferiores, dolní žíly polokoule – venae hemispheriae inferiores, dolní červová žíla – vena vermis inferior, předcentrální mozečková žíla – vena preacentralis cerebelli, skalní žíla Dandy – vena petrosa Dandy, na dolní a laterální ploše mozečku, které vtékají do příčného spalvu – sinus transversus, dolního skalního splavu – sinus petrosus interiér a esovitého splavu – sinus sigmoideus. V oblasti hrotu pyramidy dorsálně od nervus trigeminus je tvořena jedna významnější skalní žíla – vena petrosa Dandy, která je nejvýznamnějším odtokem z oblasti mozečku a kmene. Dále středomozkové žíly – venae mesencephalicae – přední mostní středomozková žíla – vena pontomesencephalis anterior, mostní žíly – venae pontis, žíly prodloužené míchy – venae medulae oblongatae, žíla bočního vychlípení čtvrté komory – vena recessus lateralis ventriculi quarti.
Žíly z prodloužené míchy a Varolova mostu jsou spojeny s žilami míchy hřbetní, zpravidla jsou tvořeny dvěmi mediánními žilami – přední a zadní. Zadní odtéká do týlního splavu – sinus occipitalis, z přední jdou větvičky podél nervů, zejména n. hypoglossus do žil v podjazykovém kanálku – canalis hypoglossi. Žíly mostu tvoří síť na basální straně, která kaudálně souvisí se žilami prodloužení míchy, kraniálně se žilami v oblasti kolem pedunculi cerebri, po stranách a vzadu pak s žilami mozečkovými.
Vnitřní hrdelní žíla – vena jugularis interna odvádí téměř všechnu krev z dutiny lebeční. Začíná na bázi lební v laterální část hrdelním otvoru – foramen jugulare rozšířením zvaným horní bulva vnitřní hradelní žíly – bulbus superior venae jugularis internae. K hrdelnímu otvoru – foramen jugulare se sbíhají nitrolební žilní splavy. Horní bulva – bulbus superior tvoří v kosti skalní – os petrosum hrdelní jámu – fossa jugularis. Horní bulva – bulbus superior naléhá ke spodině dutiny středoušní a může se do ní i vyklenovat. Vnitřní hrdelní žíla – vena jugularis interna sestupuje na krku podél vnitřní krkavice ke krkavici společné, zprava zezadu od ní a potom laterálně, a nakonec ventrolaterálně od tepny. Má průsvit 8–18.10-3 m, vlevo bývá užší. Před soutokem s podklíčkovou žílou – vena subclavia se rozšiřuje ve vřetenovitou dolní bulvu vnitřní hrdelní žíly – bulbus inferior venae jugularis internae, opatřenou jednoduchou, někdy dvojitou, chlopní, často insuficietní. Do vnitřní hrdelní žíly – vena jugularis interna vstupují žíly z lebky, ze zevní báze lební, z horní části krku, krčních orgánů, a žíly obličejové.
Lze je systémově rozdělit na přítoky.
- intrakraniální a
- extrakraniální.
Mezi intrakraniální přítoky patří:
- Splavy tvrdé pleny – sinus durae matris
- Plenové žíly – venae meningeae
- Výtokové žíly – venae emissariae
- Meziploténkové žíly – venae diploicea
- Labyrintové žíly – venae labyrinthi
- Oční žíly – venae ophthalmicae.
Plenové žíly – venae meningeae jsou úzké durální žíly provázející obdobné tepny. Nejmohutnější z nich je střední plenová žíla – vena meningea media, zpravidla dvojitá. Ústí přes trnový otvor – foramen spinosus spolu s plenovou tepnou – arteria meningica do křídlovité pleteně – plexus pterygoideus.
Výtokové žíly – venae emissariae jsou spojky, které spojují splavy žilní se žilami extrakraniálními přes otvory kostní v lebce, tzv. emissaria. Jsou to:
- temenní výtok – emissarium parietale, spojující horní šípový splav – sinus sagitalis superior se žilami měkkých pokrývek lebních,
- bradavkový výtok – emissarium mastoideum, který vede z esovitého splavu – sinus sigmoideus do týlní žily – vena occipitalis nebo do zaušní žíly – vena retroauricularis,
- týlní výtok – emissarium occipitale (poměrně vzácný), jež spojuje soutok splavů – confluens sinuum s některou z okcipitálních žil a přijímá často týlní meziploténkovou žilu – vena diploica occipitalis,
- hrbolový výtok – emissarium condylare spojuje esovitý splav – sinus sigmoideus cestou hrbolového kanálku – canalis condylaris s páteřní žilní pletení – plexus venosi vertebrales,
- žilní pleteň oválného otvoru – plexus venosus foraminis ovalis v oválném otvoru -foramen ovale spojuje dutinkový splav – sinus cavernosus do křídlovité pleteně – plexus pterygoideus,
- žilní pleteň podjazykového kanálku – plexus venosus canalis hypoglossi, žilní síť opřádá nervus hypoglossus v canalis hypoglossus a jdoucí z týlního splavu – sinus occipitalis do konečné části dolního skalního splavu – sinus petrosus inferior, před jeho vyústěním do horní bulvy hrdelní vnitřní žíly – bulbus superior venae jugularis internae,
- žilní pleteň vnitřní krkavice – plexus venosus caroticus internus opřádá vnitřní krkavici – arteria carotis interna v krkavicovém kanále – canalis caroticus,
- žilní spojky intrakraniálně extrakraniální, které se dochovaly vývojově – jsou to otvůrky na různých místech báze lební. Patří k nim spojky dutinkového splavu – sinus cavernosus (nebo circularis) přes tělo klínové kosti – corpus ossis sphenoidalis, žilky do pleteně hltanové, spojky přes tzv. canaliculus inominatus Arnoldi atd.
Meziploténkové žily – venae diploicae jsou v kanálcích diploe kostí tvořících lebeční klenbu – calvarii, v tzv. meziploténkových kanálech -canales diploici Brescheti. Probíhají vlnovitě a mají četné vtokové větve. Často překračují hranice jednotlivých kostí přes sutury, zejména pokud došlo k synostosis suturarum. Patří k nim: pravá a levá čelní meziploténkové žíly – vena diploica frontalis dextra et sinitra ústící do horního šípového splavu – sinus sagitalis superior, přední spánková meziploténková žíla – vena diploica temporalis anterior v laterální okraji šupiny čelní, která ústí do klínotemenního splavu – sinus sphenoparietalis i do hlubokých spánkových žil – venae temporales profundae. Střední spánková meziploténková žíla – vena diploica temporalis media je nekonstantní a ústí do horního skalního splavu – sinus petrosus superior. Zadní spánková meziploténková žíla – vena diploica temporalis posterior začíná v kosti temenní a ústí do příčného splavu – sinus transversus nebo přes bradavkový výtok – emissarium mastoideum. Týlní meziploténková žíla – vena diploica occipitalis ústí do příčného splavu – sinus transverus, ale i do žilních otvůrků u kondylů týlních.
Labyrintové žíly – venae labyrinthi jsou žíly odvádějící krev z vnitřního ucha a jsou celkem tři, z toho dvě intrakraniální:
- vnitřní sluchová žíla – vena auditiva interna seu labyrintová žíla – vena labyrinthi jde cestou vnitřního zvukovodu – meatus acusticus internus a otvírá se do dolního skalního splavu – sinus petrosus inferior,
- žíla předsíňového mízovodu – vena aqueductus vestibuli vede stejnojmenným kanálkem a ústí do horního skalního splavu – sinus petrosus superior,
- žíla hlemýžďového kanálku – vena canaliculi cochleae vyúsťuje extrakraniálně do konečné části dolního skalního splavu – sinus petrosus inferior nebo přímo do horní bulvy hrdelní žíly – bulbus superior venae jugularis internae.
Oční žíly – venae ophthlamicae odvádí krev z očnice. Jsou dvě:
- horní oční žíla – vena ophthalmica superior, která probíhá spolu s oční tepnou – arteria ophthalmica. Začíná na mediální straně oční koule, křižuje optický nerv směrem laterálním, prochází skrze fissura orbitalis superior a ústí do sinus cavernosus. Má soutok s analogními větvemi arteria ophthalmica. Při mediálním koutku očním je na začátku spojena s nosočelní žílaou – vena nasofrontalis a úhlovou žílou – vena angularis, a tím i s oblastí obličejové žíly – vena facialis. Přítoky žilní z koule oční jsou centrální sítnicová žíla – vena centralis retina, vírové žíly (cévnatkové) – venae vorticosae, řasové žíly – venae ciliares,
- dolní oční žíla - vena ophthalmica inferior začíná ze žilní pleteně uložené pod oční koulí a mezi přímým očním svalem – dolní a boční – musculus rectus bulbi inferior a musculus rectus bulbi lateralis. Ve svých začátcích je spojena s oblastí horní oční žíly – vena ophthalmica superior, odvádí krev skrze dolní očnicovou štěrbinou – fissura orbitalis inferior do křídlovité pleteně – plexus pterygoideus. Má spojky do hluboké obličejové žíly – vena facialis profunda.
Žilní nitrolební splavy – sinus durae matris
Splavy lební – sinus durae matris, jsou široké žilní kanály zavzaté do dura mater – tvrdé pleny mozkové, která vystýlá vnitřní plochu lebečních kostí.
Za vývoje se tvoří z žilních pletení, uložených v mesenchymu obklopujícím mozek. Teprve s diferenciací dura mater jsou do ní přijaty a přitlačeny k lebečním kostem.
Mají endoteliální výstelku a jejich stěny neobsahují hladké svalstvo, ale jen fibrilární vazivo, takže neustále zejí na průřezu. Stěna je tuhá, málo elastická, málo poddajná. V sinech chybí chlopně. Uvnitř horního šípového splavu – sinus sagitalis superior a příčného splavu – sinus transversus jsou rozepjaty tuhé vazivové trámce různého typu – chorda Willisi (Willis, 1664), kryté endotelem., V sinus cavernosus tvoří trámečky hustou síť.
Chorda Willisi (Thomas Willis, 1664) jsou struktury vazivového charakteru umístěné v lumen sinus sagitalis superior. Tyto struktury vzhledem k uložení a různým typům ovlivňují hemodynamiku odtoku krve.
Chordae Willisi dělíme na typy:
- trabekulární „podobnou chlopni“ – tzv. „valvelike“ typ akcesorní – přídatnou chordu v malém procentu případů
- akcesorní – přídatnou chordu v malém procentu případů
Žilní mozkové splavy jsou:
Nepárové:
horní šípový splav – sinus sagitalis superior,
dolní šípový splav – sinus sagitalis inferior,
přímý splav – sinus rectus,
týlní splav – sinus occipitalis,
bazilární splav – sinus basilaris.
Párové:
pravý a levý příčný splav – sinus transversus dexter et sinister,
pravý a levý esovitý splav – sinus sigmoideus dexter et sinister,
pravý a levý horní skalní splav – sinus petrosus superior dexter et sinister,
pravý a levý dolní skalní splav – sinus petrosus inferior dexter et sinister,
pravý a levý klínotemenní splav – sinus sphenoparietalis (Brescheti) dexter et sinister – je tradován – de facto neexistuje
Speciální:
dutinkový splav – sinus cavernosus a mezidutinkový splav přední a zadní – sinus intercavernosus anterior et posterior tvoří kolem hypofýzy tzv. okruh žilní – circulus venosus Ridleyi či kruhový splav – sinus circularis Ridleyi.
Přemosťující žíly mozku
Přemosťující žíly mozku jsou ty žilní struktury, které navazují na povrchové žíly mozku a vytvářejí spojení mezi jimi a nekolabovatelným žilním systémem – siny mozkovými. Tyto žíly se nacházejí v oblasti parasagitální, oboustranně falxu na konvexitě, dále na spodině mozku v oblasti pterionální(bod pterion na lebce), ve střední jámě lební v místě sinus sigmoideus a v oblasti zadní jámy lební. Je popsána pravidelně vystupující žilní struktura typu přemosťující žíly v oblasti lamina cribrosa jednostranně či oboustranně, která vstupuje do lamina cribrosa a spojuje tak extrakraniální žilní systém se žilním systémem intrakraniálním, v 15 % je vyvinuta velmi výrazně, vždy ale jednostranně, probíhá do žilního systému podél bulbus a tractus olfactorius. Procházejí arachnoidální vrstvou mezi trabekulami a navazují na povrchový žilní systém mozku, který leží pod vrstvou arachnoidální na pia mater. Jsou tenkostěnné. Některé navazují na specifické žilní systémy mozku, na anastomosy – Trolardovu horní žilní anastomosu, Labbého dolní žilní anastomosu či v oblasti kmene a mozečku je konstituována Dandyho žíla, která běží variabilně v oblasti kolem nervus trigeminus směrem k petrosnímu splavu. Je v podstatě jedna z nejdůležitějších odtokových systémů kmene a mozečku.
V oblasti kolem kmene a mozečku se nacházejí četné žilní spojky do oblasti splavů na konvexitě mozečku (horní ploše, vermis, laterálních plochách), kolem hlavových nervů (postranního smíšeného systému, nervus hypoglossus) a spolu s některou vertebrální arterií, vzácněji jde přemosťující žíla v oblasti přes velkou cisternu (asi v 7–10 %).
Mezi přemosťující žíly tak patří i velká mozková žíla – vena magna cerebri Galleni.
Z povrchových žil jdou přemosťující žíly: ze dvou prefrontálních povrchových mediálních horních (venae prefrontales superficiales medii superiores – 3–5), po jedné z předcentrální horní povrchová střední žíla – vena precentralis superficalis medii superior, žíla čichovéo závitu – vena gyry olfactorii, přední mozková žíla – vena cerebri anterior, a dále žíly do dolního šípového splavu – sinus sagitalis inferior 3–5, 1–3 z horní centrální povrchové střední žíla - vena centralis superficialis media superior, 2–3 z horní střední temenní povrchové žíly - venae parietales superficialies medii superiores a střední povrchní týlní žíly – venae occipitales superficiales medii.
| Obrázky ke kapitole: Cévní zásobení centrálního nervstva (anatomie cévního systému mozku) | |||

Obr. 64 Tepny na bazi mozkové
|

Obr. 65 Přední a zadní mozková tepna
|
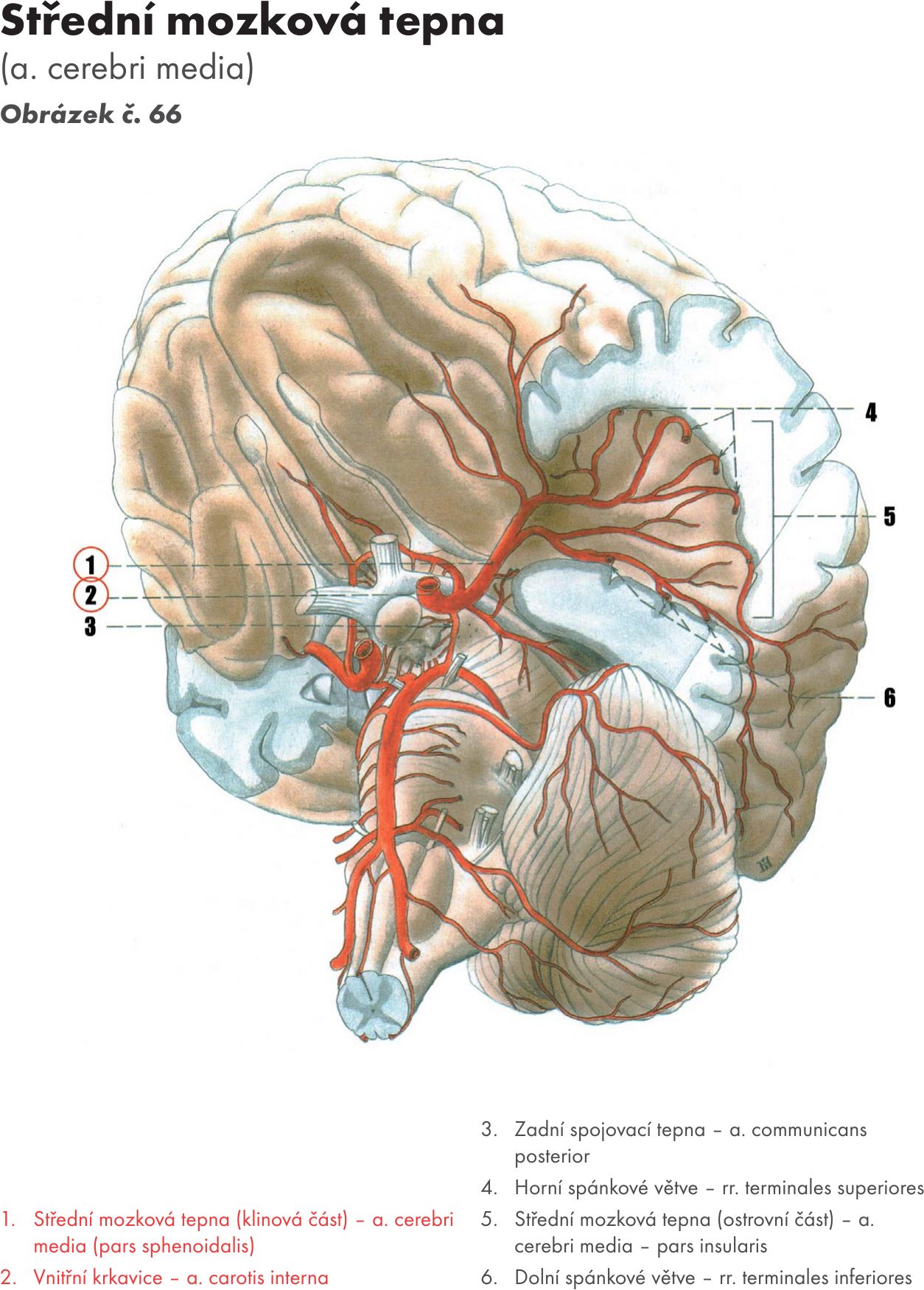
Obr. 66 Střední mozková tepna
|
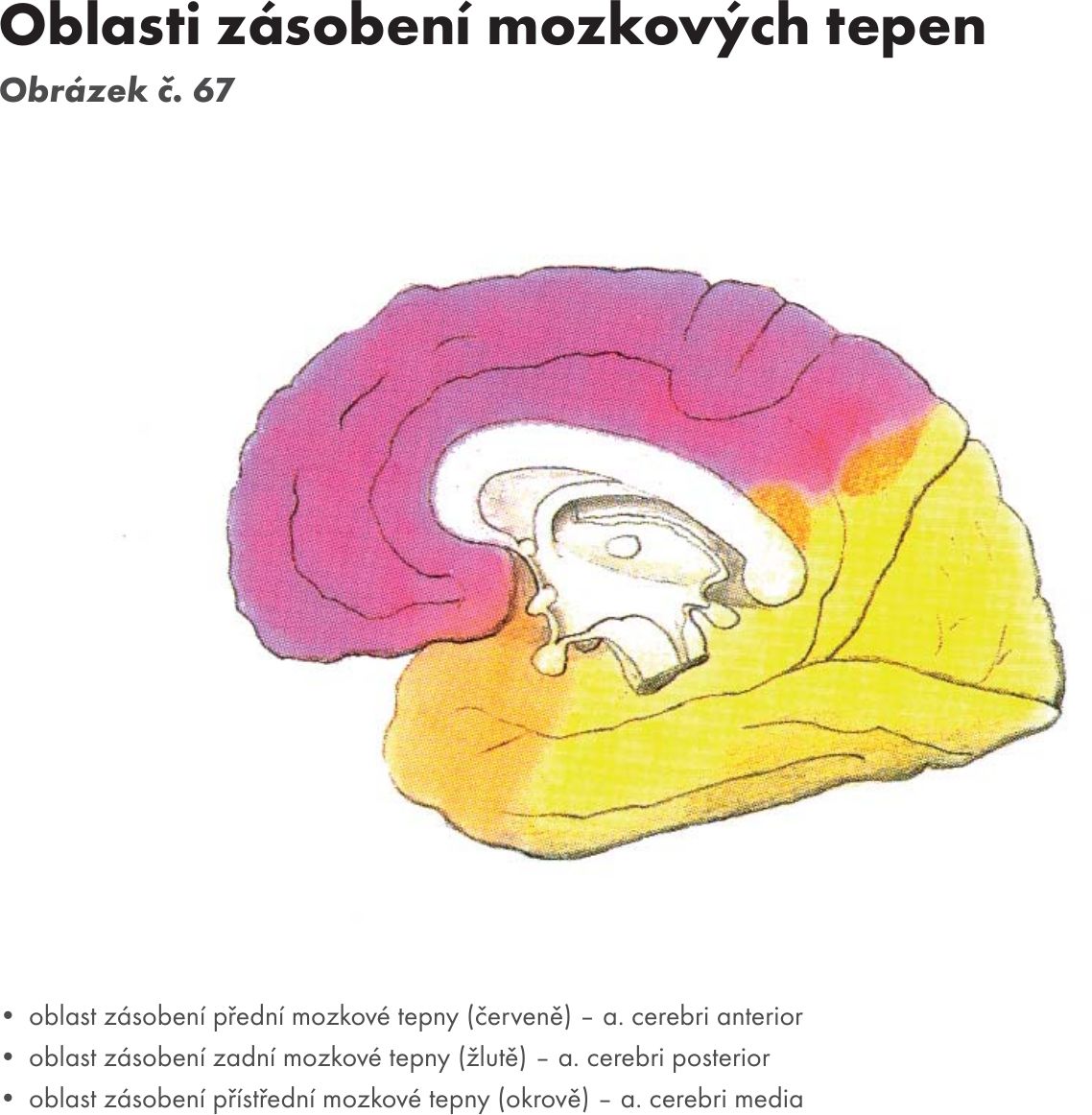
Obr. 67 Oblasti zásobení mozkových tepen
|
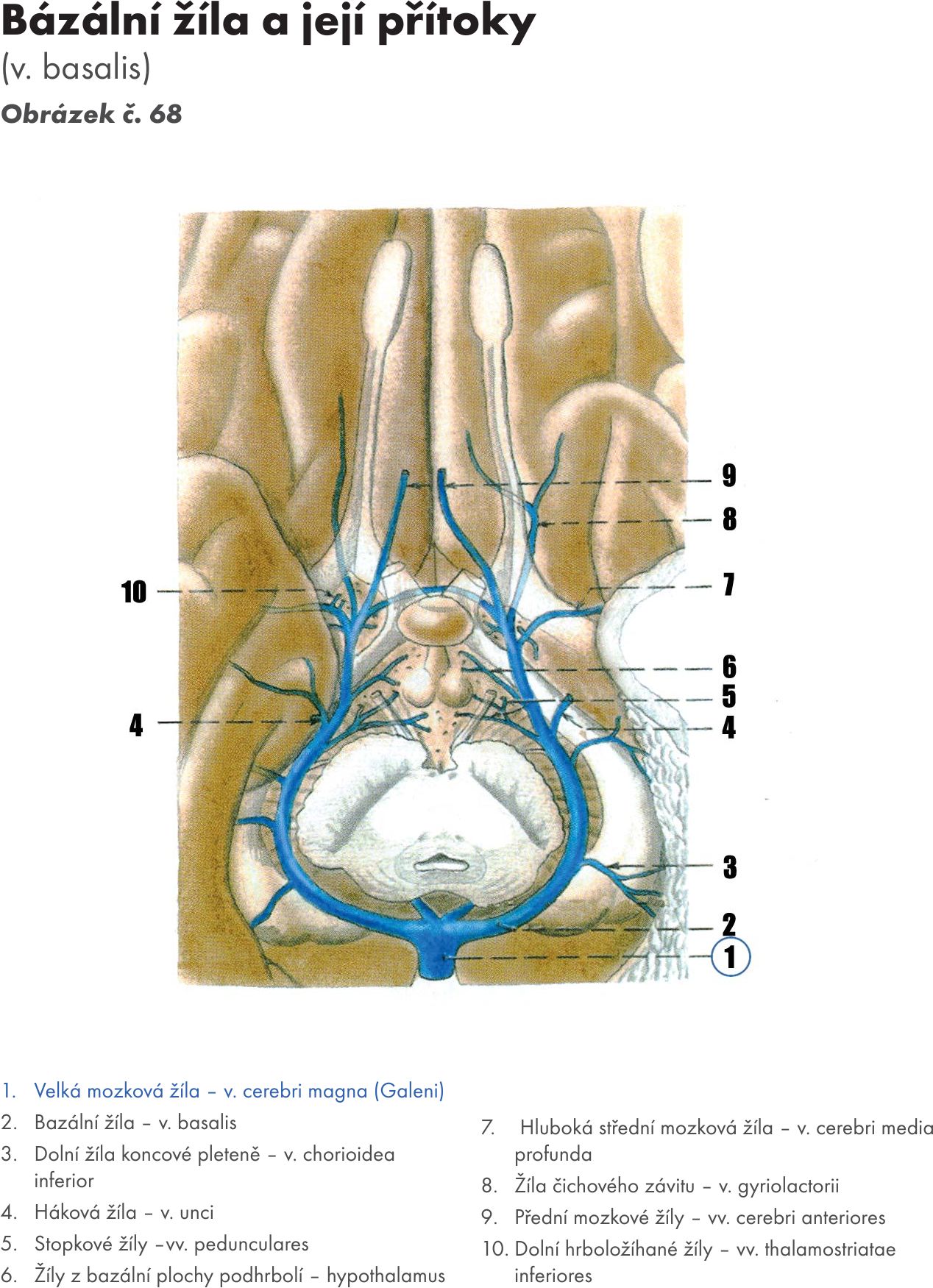
Obr. 68 Bazální žíla a její přítoky
|

Obr. 69 Hluboké mozkové žíly
|
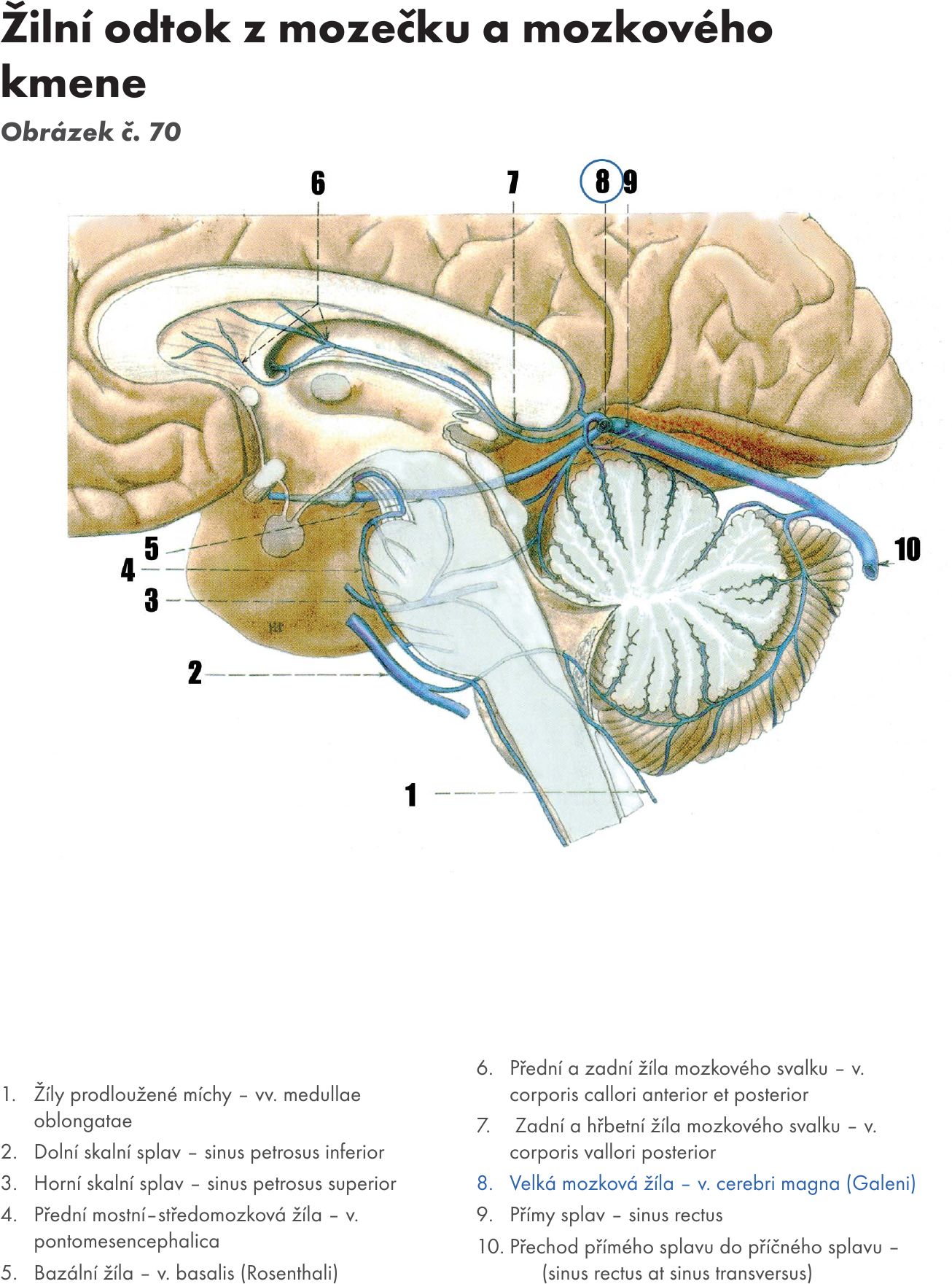
Obr. 70 Žilní odtok z mozečku a mozkového kmene
|
|
Spoje centrálního nervového systému
-
Motorické dráhy
Slouží k převádění výstupních informací CNS k efektorům, dělíme je na somatomotorické a visceromotorické.
Činnost kosterního svalstva zajišťuje:
- Mimovolní pohyby řízené reflexně (vzpřímená poloha, obrana organismu…)
- Volní pohyby (pohyb při pracovních výkonech, řeči, volní hybnost…)
Pohybové funkce jsou řízeny podle fylogenetického řádu:
- Nejjednodušší reflexy řídí míšní aparát
- Nadřazené řídicí systémy jsou v mozkovém kmeni (F.R.), podkorových oblastech a kůře mozkové
Vyšší oddíly jsou funkčně nadřazeny nižším, jejich činnost je harmonicky skloubena a všechny jejich sestupné (descendentní) dráhy končí na posledních motorických neuronech míchy – ncll. motorii a motorických jádrech hlavových nervů- ncll. originis n. III., IV., V., VI., VII., IX., X., XI., XII., které jsou konečnou společnou dráhou motorických systémů.
Pohyb kosterního svalstva realizují:
- Pyramidový systém
- Extrapyramidový systém
- Mozečkový systém
Pyramidové dráhy
Obrázek č. 73 Obrázek č. 74 Obrázek č. 75
Označují se též jako přímé motorické dráhy. Spojují motorická kortikální centra jedním neuronem s motorickými jádry míšních a hlavových nervů. Pyramidové dráhy jsou fylogeneticky nejmladší a u člověka jsou mohutně vyvinuty. Vlákna myelinisují až postnatálně ve 2.–4. roce života. Pyramidové dráhy začínají především v předcentrálním závitu – gyrus precentralis (area 4) na velkých pyramidových Běcových buňkách. Jejich neurity tvoří 2–3 % pyramidových drah. Buňky zachovávají přesné somatotropické uspořádání, ale tělo se do gyru promítá postavením vzhůru nohama
Přímé korové dráhy se dělí na:
- Koromíšní dráhu – tractus corticospinalis
- Korojádrovou dráhu – tractus corticonuclearis
Koromíšní dráha - tractus corticospinalis, je jednoneuronová dráha, která začíná v senzomotorických korových oblastech. Sestupuje skrze zadní raménko vnitřního pouzdra-crus posterius capsulae internae. Má somatotopické uspořádání, vlákna pro HK jdou vepředu, vlákna pro DK vzadu. Dále prochází ve střední části mozkových ramen středního mozku-crura ceribri mesencephala, pokračuje do mostu, kde se tříští o jeho jádra-nuclei pontis.
Na přední ploše prodloužené míchy tvoří hrbolky – pyramis medullae oblongatae. Pod pyramidami se dráha zčásti kříží-decussatio pyramidum. Její zkřížená část pokračuje jako boční koromíšní dráha-tr. corticospinalis lateralis (80 % vláken) do postranního provazce míšního, nezkřížená část pokračuje jako přední koromíšní dráha – tr. corticospinalis anterior (20 % vláken) do ventrálního provazce míšního. Část těchto vláken se však též kříží v bílé spojce - commisura alba. Dráha končí na motoneuronech předních rohů míšních, interneuronech a neuronech baze zadních rohů (kontrolní systém).
Korojádrová dráha - tractus corticonuclearis, neurony končí na motorických jádrech (nuclei originis) hlavových nervů. Pro n. trigeminus (n. V.) žvýkací svaly, n. facialis (n. VII.) mimické svaly, n. glossopharyngeus (n. IX.) svaly patra a pharyngu, n. vagus (n. X.) svaly pharyngu a laryngu, n. accesorius (n. XI.) m. sternocleidomastoideus a trapezius a n. hypoglossus (n. XII.) svaly jazyka. Začínají převážně v :
- Gyrus preacentralis (area 4)
- Gyrus frontalis superior et medius (area 8)
- Gyrus frontalis inferior (area 44)
Pro motorická jádra okohybných nervů: n. oculomotorius (n. III.), n. trochlearis (n. IV.) a n. abducens (n. VI.) začínají neurony převážně v
- Gyrus preacentralis (area 4)
- Gyrus occipitalis (area 19)
Dráha sestupuje v koleně vnitřního pouzdra – genu capsulae internae, do středního mozku (mesencefala), kde lemuje mediálně i laterálně v crus cerebri tr. cerebrospinalis. Tak jako v mesencefalu i v nižších etážích kmenu mozkového – mostu (pontu) a prodloužené míše (medulla oblongata), končí vlákna na příslušných jádrech hlavových nervů. Vlákna tractus corticonuclearis se za průběhu mostem a prodlouženou míchou částečně kříží. Většina jader hlavových nervů dostává vlákna zkřížená i nezkřížená.
Extrapyramidové dráhy Obrázek č. 76
Označují se též jako nepřímé motorické dráhy. Spojují mimopyramidové oblasti kůry mozkové s posledními motorickými neurony podkorových a kmenových jader. Dráhy jsou víceneuronové a mají 3 úseky
- Spojení kůry s basálními ganglii a jádry kmene
- Spojení basálních ganglií
- Spojení kmenových jader s posledními motorickými neurony
- Spojení kůry s basáními ganglii a jádry kmene (kortikální systém)
- Dráha koroproužková – tr. corticostriatici: spojuje neocortex (area 4,6,8,9,12,19) s ocasatým jádrem – ncl. caudatus a area 4 se skořápkou – putamen. (striatum=ncl. caudatus+putamen)
- Dráha korobledá – tr. corticopallidalis: spojuje area 6 s bledou kulovitou hmotou (jádro) - globus pallidus
- Dráha koročervená – tr. corticorubralis: spojuje kůru gyri orbitales, gyrus prae i postcentralis a gyri frontales s ncl. ruber
- Dráha koročerná – tr. corticonigralis: spojuje area 4 a 6 s černou hmotou – substantia nigra
- Dráha koropodhrbolová – tr. corticosubthalamicus: spojuje area 4 a 6 s podhrbolovým jádrem – ncl. subthalamicus (Luysi)
- Dráha korokrytová – tr. corticotectalis: spojuje area 19 se středomozkovým krytem – tectum mesencephali
- Dráha koropokrývková– tr. corticotegmentalis: spojuje area 4 a 6 se středomozkovou pokrývkou – tegmentum mesencephli
- Dráha korosíťová – tr. corticoreticularis: spojuje area 4, 8, 24, 2, 19 s jádry F.R. (síťovaná formace – formatio reticularis)
Další dráhy spojují mozek s mozečkem (neocortex s neocerebelární kůrou mozečku)
- Dráhy koromozečkové – trr. corticocerebellares: spojují kůru motorickou, ale i senzitivní, sluchovou a zrakovou s mostními jádry – nuclei pontis. Zde se dráhy přepojují a tím se rozpadají na dráhu koromostní – tr. corticopontinus, a dráhu mostomozečkovou – tr. pontocerebellaris (tr. cortico-ponto-cerebellaris). Do mozečku jdou přes střední mozečkovou stopku – pedunculus cerebellaris medalis.
Dráhy se nazývají podle toho, z kterého mozkového laloku vycházejí:
- Dráha čelněmostní – tr. frontopontinus (Arnold) : Začíná v čelní kůře – lobus frontalis, v area 4 a 6. V capsula interna probíhá v crus anterius, ve středním mozku nejmediálněji v crus cerebri, v mostu se přpojuje na ncll. pontii a zkříženě pokračují do kůry mozečku (neocerebellární kůry).
- Dráha spánkomostní – tr. temporopontinus (Türck): Začíná ve všech spánkových závitech – gyrus temporalis superior, medius et inferior. V capsula interna probíhá v crus posterius, ve středním mozku nejlaterálněji v crus cerebri, v mostu se přepojuje na ncll. pontis a zkříženě pokračuje do neocerebellarní kůry.
K této dráze se ještě přidávají: - Dráha temenomostní – tr. parietopontinus : (area 3, 2, 1, 5)
- Dráha týlněmostní – tr. occipitopontinus: (area 19, 18, 17)
- Spojení basálních ganglií (systém striatopallidální)
Aferentní dráhy basálních ganglií přicházejí:
- Z neocortexu (trr. corticostriatici et corticopalliadales)
- Z thalamu (tr. thalamostriatici et thalamopallidales)
Striatum a pallidum spojují trr. striatopallidales
Eferentní dráhy vycházejí:
- Ze striata
- tr. striatonigralis
- Z pallida
- tr. pallidohypothalamicus
- tr. pallidosubthalamicus
- tr. pallidorubralis
- tr. pallidonigralis
- tr. pallidoreticularis
Umístění perikaryonů a zakončení neuritů ukazují názvy. Descendentně probíhající eferentní dráhy, které leží ve Forelově poli (H2, H1)
- Spojení kmenových jader s posledními motorickými neurony – nuclei motorii (truncospinální systém)
Tyto spoje navazují na mimopyramidové neurony přicházející z kůry mozkové a z bazálních ganglií. Umístění perikaryonů a zakončení neuritů ukazují názvy.
- Dráha stíťomíšní - tr. reticulospinalis, začíná ve všech systémech FR, je to dráha víceneuronová se zkříženými i nezkříženými vlákny. Je to hlavní mimopyramidová dráha u čověka a ovlivňuje řízení svalového tonu.
- Dráha červenomíšní - tr. rubrospinalis (Monakovova dráha), začíná na velkobuňečné části nucleus ruber. Vlákna dráhy se postupně kříží. Má excitační vliv na motoneurony flexorů. Přivádí vlivy nejen korové, ale i mozečkové. Má vliv na udržování svalového tonu.
- Dráha centrální pokrývková (čepcová) – tr. tegmentalis centralis, obsahuje vlákna rozmanitého původu. Jsou to většinou spoje FR. Řadíme sem dráhu červenošíťovou – tractus rubroreticularis a dráhu červenoolivovou – tractus rubroolivaris.
- Dráha olivomíšní- tr. olivospinalis, jde z olivárního jádra prodloužené míchy sestupuje v laterálním provazci (Helwegova dráha) na ncll. motorii. Pomáhá udržovat rovnováhu těla.
- Dráha krytomíšní – tr. tectospinalis, je to fylogeneticky stará dráha. Začátek je v hlubokých vrstvách colliculus superior, kříží se ještě v mesencephalu (Meynertovo křížení), sestupuje v přidaném provazci a končí na prvních krčních motorických jádrech. Slouží k opticko-motorickým a akusticko-motorickým reflexům hlavy a krku.
-
Dráhy mozečku
Obrázek č. 77 Obrázek č. 78 Obrázek č. 79Dráhy mozečku zahrnují všechny tři vývojové systémy: vestibulární (archicerebellum), spinální (paleocerebellum), cerebrální (neocerebellum) a končí vždy v příšlušné mozečkové kůře.
Senzitivní dráhy, které jdou do mozečku se nazývají nepřímé:
- Dráha přední míchomozečková – tr. spinocerebellaris anterior
- Dráha zadní míchomozečková – tr. spinocerebellaris posterior
- Dráha trojklannomozečková – tr. trigeminocerebellaris
- Dráha jádromozečková – tr. nucleocerebellaris
- Dráha bulvomozečková – tr. bulbocereberalis
Eferentní dráhy mozečku jsou někdy přímo řazeny k extrapyramidovému systému, protože mozeček představuje pomocné zařízení motorických systémů.
Dráhy mozečku uvádíme souhrnně podle původu jednotlivých částí.
- Dráhy vločkouzlíkového laloku – lobus flocculonodularis
Afferentní dráhy
- Dráha předsíňomozečková – tr. vestibulocerebellaris, vychází z terminálních jader nervi vestibuli (n.VIII). Do mozečku vstupuje přes dolní mozečkovou stopku – pendunculus cerebellaris inferior, kde končí jako mechová vlákna.
Efferentní dráhy
- Dráha uzlíkopředsíňová – tr. nodulovestibularis
- Dráha vločkopředsíňová – tr. flocculovestibularis
- Dráha štítopředsíňová – tr. fastigiovestibularis (Russelův svazek)
Dráhy končí v Deitersově vestibulárním jádře.
- Dráhy předního a zadního laloku – lobus cranialis et caudalis
Afferentní dráhy
- Dráha zadní míchomozečková – tr. spinocerebellaris posterior (Flechsigův svazek), vychází ze Stilligovo-Clarkeova jádra a do mozečku vede nezkřížená vlákna přes dolní mozečkovou stopku – pendunculus cerebellaris inferior
- Dráha přední míchomozečková – tr. spinocerebellaris anterior (Gowersův svazek), vychází ze Stillingovo-Clarkeova jádra a do mozečku vede zkřížená vlákna přes horní mozečkovou stopku – pendunculus cerebellaris superior
- Dráha bulvomozečková – tr. bulbocerebellaris, vychází z jader zadních provazů míšních (ncl. gracilis et ncl. cuneatus) a do mozečku jdou nezkříženě zadními vnějšími obloukovitými vlákny – fibrae arcuatae externae dorsales, nebo zkříženě předními vnějšími obloukovitými vlákny – fibrae arcuatae externae ventrales. Do mozečku vstupuje přes dolní mozečkovou stopku – pendunculus cerebellaris inferior.
- Dráha jádromozečková – tr. nucleocerebellaris, vychází u terminálních jader senzitivních hlavových nervů (V., VII., IX., X.). Do mozečku jsou přes dolní mozečkovou stopku – pendunculus cerebellaris inferior a přidružují se k drahám předešlým.
- Dráha olivomozečková – tr. olivocerebellaris, vychází ze starší části olivárních jader prodloužené míchy a do mozečku vstupuje přes dolní mozečkovou stopku – pendunculus cerebellaris inferior
- Dráha krytomozečková – tr. tectocerebellaris, vychází z jader pod čtverhrbolím středního mozku (tectum mesencephali) pod ncl. colliculi superiores (rostrálními hrbolky) a do mozečku vstupuje přes horní mozečkovou stopku – pendunculus cerebellaris superior, kde se kříží.
- Dráha síťomozečková – tr.reticulocerebellaris, vychází z laterálních jader FR a do mozečku vstupuje přes dolní mozečkovou stopku – pendunculus cerebellaris inferior. Dráha se nekříží.
- Dráha červenomozečková – tr. rubrocerebellaris, vychází z červeného jádra středního mozku – ncl. ruber mesencephali a do mozečku vstupuje přes horní mozečkovou stopku – pendunculus cerebellaris superior. Dráha se kříží a odbočuje z tr. rubrospinalis (Brodal)
Efferentní dráhy
Vlastní efferentní neurony mozečku mají perikaryon v mozečkových jádrech a neurit opouští mozeček k místu určení
- Dráha mozečkopokrývková – tr. cerebellotegmentalis, vychází ze zubatého jádra – ncl. dentatus přes horní mozečkovou stopku – pendunculus cerebellaris superior a končí na jádrech RF středního mozku a pontu.
- Dráha mozečkokrytová – tr. cerebellotectalis, vychází z mozečkových jader přes horní mozečkovou stopku – pendunculus cerebellaris superior do tecta.
- Dráha mozečkohrbolová– tr. cerebellothalamicus, vychází z mozečkových jader přes horní mozečkovou stopku – pendunculus cerebellaris superior k centrálním jádrům thalamu.
- Dráha mozečkoolivová – tr. cerebelloolivaris, vychází z mozečkových jader přes dolní mozečkovou stopku – pendunculus cerebellaris inferior do straší části hlavního olivárního jádra.
- Dráhy středního laloku – lobus medius
Afferentní dráhy
- Dráha mozečkomostová – tr. pontocerebellaris, vychází z jader mostu přes střední mozečkovou stopku – pendunculus cerebellaris medialis. Dráha se kříží a je pokračováním dráhy koromostové. Zajišťuje zkřížené spojení hemisfér předního mozku s hemisférami mozečku.
- Dráha olivomozečková – tr. olivocerebellaris, vychází z mladší části olivárních jader (neoolivy) prodloužené míchy a do mozečku vstupuje přes dolní mozečkovou stopku – pendunculus cerebellaris interiér.
- Dráha mozkomozečková – tr. corticocerebellaris, vychází z motorické oblasti čelního laloku a do mozečku vstupuje přes dolní mozečkovou stopku – pendunculus cerebellaris inferior. Dráhy se nekříží
Efferentní dráhy
- Dráha mozečkočervená – tr. cerebellorubralis, vychází ze zubatého jádra – nucleus dentatus a přes horní mozečkovou stopku, kde se kříží, vede do malobuněčné části nucleus ruber. Přidružuje se ke stejnojmenným drahám paleocerebella
- Dráha mozečkohrbolová – tr. cerebellothalamicus, vychází ze zubatého jádra – ncl. dentatus a přes horní mozečkovou stopku – pendunculus cerebellaris superior vede do ventrolaterálního jádra thalamu. Pokračuje jako tr. thalamocorticalis do motorické kůry mozkové (area 4,6).
-
Senzitivní dráhy
Obrázek č. 80 Obrázek č. 81 Obrázek č. 82 Obrázek č. 83Senzitivní dráhy zprostředkovávají registraci a vedení senzitivních podnětů z vnitřního a zevního prostředí až do místa jejich zpracování v CNS.
Senzitivní dráhy vedou z periferních receptorů (extero- i interoreceptorů) hmat, tlak, tah, propriocepci ze svalů a šlach, vibrace, teplo, chlad a bolest. Dělí se na dráhy protopatického hrubého a epikritického jemného čití.
- Dráhy protopatické (nociceptivní) – jejich neurony jsou fylogeneticky staré a málo myelinizované. Vedou informace nediferencované a vyvolané hrubými stimuly signalizujícími nebezpečí (bolest, teplo, chlad, hrubá kožní citlivost).
- Dráhy epikritické (diskriminační) – jejich neurony jsou fylogeneticky mladé s hodně myelinizovanými axony. Vedou informace jemně kvalitativně diferencované.
Senzitivní dráhy jsou většinou víceneuronové:
- senzitivní neuron (protoneuron) je lokalizovaný v pseudounipolárních buňkách ve spinálním gangliu míšního nervu či v příslušném sensitivním gangliu hlavového nervu.
- senzitivní neuron (deuteroneuron). Perikaryony leží v jádrech zadních rohů míšních (vlastní jádro – ncl. proprius, ncl. thoracicus (j. Stillingovo – Clarkovo) – protopatické čití, nebo v jádrech M.O. ncl. gracilis (štíhlý) et ncl. cuneatus (klínový), nebo v terminálních jádrech hlavových nervů – epikritické čití. Druhé senzitivní neurony převádějí převzaté senzitivní informace:
- do thalamu (podkoří senzitivních informací) nejkratší cestou do mozkové kůry. Tyto dráhy označujeme jako přímé senzitivní dráhy, tvoří tak zvaný lemniskový systém
- do mozečkové kůry – nepřímé senzitivní dráhy
- senzitivní neuron má své perikaryony ve specifických jádrech thalamu a jeho neurity pokračují jako projekční dráha do senzitivního asociačního centra v kůře (are 3,2,1), kde jsou lokalizovány jednotlivé vjemy.
Všechny senzitivní dráhy probíhají zkříženě.
A Přímé dráhy protopatického čití
Neurity 2. neuronu vycházejí z vlastního jádra – ncl. proprius. Konec (cíl) je patrný z názvu dráhy.
- dráha míchohrbolová zevní a přední (tr. spinothalamicus lateralis et anterior) má topické uspořádání (krční vlákna běží mediálně, lumbální laterálně) a vnitřní topografii (ve ventrodorzálním laterálním pořadí: tlak, dotek, bolest, teplota). Vede ostrou lokalizovanou bolest, chlad a teplo z opačné poloviny těla a v omezeném rozsahu také podněty hmatové. Tr. spinothalamicus anterior je převážně hmatová dráha, tr. spinothalamicus lateralis je spíše dráha bolesti a tepla.
- dráha míchosíťovaná - tr. spinoreticularis. Dráha končí na jádrech FR ležících ve třech úrovních kmene mozkového. Část retikulárních drah končí u kmenových jader, jež mají spojení do mozečku a slouží mozečkové aferentaci. Dráha vede pomalou tupou a špatně lokalizovanou difúzní bolest. Podněty z této dráhy využívá i aktivační systém FR.
- dráha míchokrytová – tr. spinotectalis. Dráha končí na jádrech v hlubokých vrstvách tekta. Dráha přenáší taktilní a nocicepční podněty do motorických systémů ovlivňujících pohyby hlavy a krku (thalamus, BG, černá hmota – substantia nigra. Z těchto útvarů se podněty sestupnými drahami (tr. tectospinalis, tr. tectoraticularis, tr, tectonuclearis) vracejí zpět k centrům, které přímo vykonávají motoriku míšní a motoriku hlavových nervů s koordinací zrakových podnětů.
B Přímé dráhy epikritického čití
Neurity 2. neuronu vychází z ncl. gracilis (štíhlý) a ncl. cuneatus (klínový).
- Ascendentní kolaterály se po překřížení – decussatio lemniscorum zatačejí nahoru – dráha bulvohrbolová – tr. bulbothalamicus
- Descendentní kolaterály přicházejí do míchy krční, hrudní a bederní – dráha míchobulvová – tr. spinobulbaris.
Přímé dráhy epikritického a protopatického čití se sdružují v objemnou prostřední kličku (snopec) – lemnicus medialis (sensitivus), který končí na specifických jádrech thalamu. Tento snopec sdružuje dráhy: tr. bulbothalamicus, tr. spinothalamicus a připojuje se tr. trigeminothalamicus.
C Nepřímé dráhy protopatického čití
Vedou informace proprioceptivní, exteroceptivní a částečně i interoceptivní do kůry mozečku. Končí hlavně v kůře spinálního mozečku (palaeocerebellum), kde jsou zpracovány pro funkci mozečku. Neurity 2. neuronu vycházejí z jádra Stillingovo – Clarkova dvojím způsobem:
- dráha míchomozečková zadní – tr. spinocerebellaris posterior. Vystupuje nahoru na stejné straně v bočním provazci – funiculus lateralis a přes ocasní mozečkovou stopku – pedunculus cerebellaris inferior končí v kůře lobus inferior cerebelli.
- dráha míchomozečková přední – tr. spinocerebellaris anterior. Dráha kříží střední rovinu, vystupuje až do středního mozku (mesencephalu) a cestou hlavovou mozečkovou stopkou – pedunculus cerebellaris superior končí v kůře lobus superior cerebelli.
D Nepřímé dráhy epikritického čití
Vedou informace proprioceptivní i exteroceptivní. Neurity 2. neuronu vycházejí z ncl. gracilis a ncl. cuneatus do mozečku a vytvářejí:
- dráhu bulvomozečkovou – tr. bulbocerebellaris. Neurity jdou dvojí cestou:
- některé neurity jdou na opačnou stranu a po překřížení jdou po povrchu olivy jako břišní vnější oblouková vlákna – fibrae arcuatae ext. ventralis
- jiné neurity jdou po své straně jako hřbetní vnější oblouková vlákna – fibrae arcuatae ext. dorsalis. Nekříží se.
Obojí neurity pokračují cestou dolní mozečkové stopky – pedunculus cerebellaris inferior do mozečku
Nepřímé dráhy protopatického i epikritického čití pokračují z mozečku opět do podkorového senzitivního centra jako dráha mozečkohrbolová – tr. cerebellothalamicus.
E Senzitivní dráhy hlavových nervů
Senzitivita je vedena vlákny hlavových nervů (V., VII., IX., X.). Funkčně tvoří smíšenou dráhu protopatické i epikritické citlivosti. Jde o citlivost z oblasti obličeje a přední části hlavy. Somatosenzitivní i viscerosenzitivní vlákna těchto nervů jsou přepojována v terminálních jádrech n. trigeminus (V.). Tato jsou analogií jádrům hlavních senzitivních míšních drah. Z jader n. trigeminus zprostředkovávají vedení 4 dráhy:
- dráha trojklanně hrbolokorová – tr. trigeminothalamocorticalis
- dráha trojklanně hrbolohrbolová – tr. trigeminothalamothalamicus
- dráha trojklanně hrbolosíťovaná - tr. trigeminothalamoreticularis
- dráha trojklanně hrbolokrytová – tr. trigeminothalamotectalis
Dráhy probíhající do specifických jader thalamu se kříží a tvoří lemniscus trigeminalis, který se připojuje k kličce (snopci) prostřednímu (cítivý) – lemniscus medialis (sensitivus). Část neuronů probíhá jako nepřímé senzitivní dráhy do mozečku. Ze specifických jader thalamu pokračuje 3. neuron, jako u předchozích drah – tr. thalamocorticalis do senzitivních asociačních center kůry (area 3,2,1)
-
Chuťová dráha
Obrázek č. 84Je soubor neuronů, které převádějí chuťové informace na různá místa CNS a do mozkové kůry. Projekční neurony do kůry umožňují vnímání chuti.
Orgánem chuti jsou chuťové pohárky – caliculi gustatorii, které jsou uloženy především ve sliznici jazyka, patra a zadní stěně hltanu. Hlavní součástí chuťových pohárků jsou sekundární smyslové chuťové buňky. Dráha má tři neurony:
- 1. neuron
- spojuje různými cestami chuťové buňky (které opřádá svými dendrity) s jádrem osamocené dráhy – ncl. tractus solitarii. Spojení se uskutečňuje cestou lícního nervu – n. facialis (n. VII), jeho smíšenou částí – prostřední lícní nerv – n. intermediofacialis, cestou jazykohltanového nervu – n. glossopharyngeus (n. IX) a cestou nervu bloudivého – n. vagus (n.X)
- vlákna z prostředního lícního nervu – n. intermediofacialis převádějí chuťové informace z předních 2/3 jazyka (část inervovaní jazykovým nervem – n. lingualis (z n. V). Pseudounipolární buňky leží v kolínkové nervové uzlině – ganglion geniculi. Periferní výběžky přicházejí od chuťových buněk nejprve ve svazku n. lingualis, potom v bubínkové struně – chorda tympani a dále se přidávají k n. VII a vstupují do jednotné dráhy osamocené – tractus solitarius
- periferní výběžky v patrových nervech – nn. palatini přivádějí informace z měkkého patra, prochází přes křídlopatrovou nervovou uzlinu – ganglion pterygopalatinum a cestou velkého skalního nervu – n. petrosus major se přidávají k n. VII a vstupují do dráhy osamocené – tractus solitarius
- vlákna n. glossopharyngeus (n. IX) přivádějí informace ze zadní 1/3 jazyka a oblasti hrdelní úžiny – isthmus faucium. Dendrity vedou do ganglií n. IX a po přepojení vstupují kmenem n. IX do tractus solitarius
- vlákna n. vagus (n. X) přivádějí chuťové informace z okolí epiglottis. Dendrity vedou do ganglií n. X a po přepojení vstupují kmenem n. X do dráhy osamocené - tractus solitarius
- 2. neuron
- Neurony jsou lokalizovány v jádru dráhy osamocené – ncl. tractus solitarius, jejich axony běží cestou prostřední kličky (snopce) – lemniscus medialis a končí jako dráha osamocenohrbolová – tractus solitariothalamicus ve specifických jádrech mezimozkového hrbolu (thalamu). Kříží střední rovinu a přidávají se k ostatním senzitivním drahám. Část druhých neuritů se větví v okolí motorických jader hlavových nervů a slouží pro reflexní oblouky. Jiné se napojují na síťovanou formaci – formatio reticularis (FR) a končí v krční míše.
- 3. neuron
- Projekčni neurony spojují specifický hrbol – thalamus s korovým chuťovým centrem. Otázka chuťového projekčního centra není přesně vyřešena, pravděpodobná lokalizace je v dolní části zacentrálního závitu – gyrus postcentralis (area 43). Jako další možnost se uvádí: hák příhippokampového závitu – uncus gyri parahippocampalis a přední části ostrova – insuly. K jemnější chuťové diskriminaci je potřeba součinnosti chuťové a čichové dráhy dohromady. (hippocampus = mořský koník)
- 1. neuron
-
Čichová dráha
Představuje soubor neuronů, které převádějí čichové podněty z čichové části nosní sliznice – regio olfactoria na různá místa CNS, kde jsou zpracovány a zužitkovány. Čichová dráha je dvouneuronová.
- 1. neuron
- Představují primární bipolární čichové buňky uložené v regio olfactoria nosní sliznice (viz Základy anatomie III, s. 10). Buňky jsou schopny transportovat čichové vjemy na nervové impulzy. Periferní krátké výběžky tvoří dendritickou zónu. Neurity těchto buněk tvoří čichové nervy – nervi olfactorii, které prostupují stropem dutiny nosní – lamina cribrosa ossis ethmoidalis do čichové bulvy – bulbus olfactorius, kde se rozpadají na jemné větévky. Oba bulbi olfactorii jsou navzájem propojeny komisurálními neurony, které se kříží přes přední část – commissura anterior. Do bulbus olfactorius přicházejí též eferentní neurony mozkové kůry, které mají regulační vliv.
- 2. neuron
- Představují mitrální buňky v bulbus olfactorius. Jejich periferní výběžek se rozvětvuje v dendritickou zónu ve formě krátkých větévek, které navazují na podobné rozvětvení prvního čichového neuronu, čímž vzniká klubíčkovitý útvar – glomerulus olfactorius. Neurity mitrálních buněk tvoří čichovou dráhu – tractus olfactorius, která končí v různých (převážně starých – palaeokorových) oblastech mozkové kůry. Za čichový korový analyzátor je považován např.: hák přihipokampového závitu – uncus gyrii parahippocampalis nebo mandlové jádro – ncl. amygdaloideus. Eferentní neurony palaecortexu představují aferentní vstupy do dalších systémů CNS (do limbického systému, F.R., hypothalamu, thalamu)
Funkce čichové dráhy
Receptory čichových buněk přijímají chemické signály ze zevního prostředí, které jsou postupně přeneseny do čichových korových oblastí. Napojením čichových drah na limbický systém, hypothalamus a na další struktury vzniká současně emoční odpověď na čichové podněty. Napojení na autonomní nervový systém vyvolává visceromotorické odpovědi (viz dráhy limbického systému – str.).
- 1. neuron
-
Mediátory CNS a chemické systémy
Akční potenciál vyvolává uvolnění mediátoru (neurotransmiteru), což je základem přenosu vzruchů v synapsích. Mediártor se váže na receptor na postsynaptické membráně. Receptorem je protein, jeho chemickoprostorové uspořádání, membrány nervové či jiné buňky efektoru (svalové, žlázy apod.).
Klasické, rychle působící mediátory jsou malé molekuly (aminy aminokyseliny). Spolu s klasickými mediátory se vyskytují mediátory s velkou molekulou (neuropeptidy) většinou s pomalým a dlouhodobým účinkem. Neuromodulátory jeou chemické látky (často peptidy) nacházející se v synapsích současně s dalšími mediátory a ovlivňující synaptický přenos.
Ke klasickým mediátorům patří : acetylcholin, katecholaminy (adrenalin, noradrenalin, dopamin), indolaminy(serotonin, histamin) aminokyseliny, peptidy - substance P, enkefaliny, endorfiny; plyny – CO,NO.
V CNS rozlišujeme různé systémy dle mediátorů:
- Cholinergní systém - acetylcholin nemá jednotnou funkci, je mediátorem motorických a vegetativních funkcí, učení a paměti. Dle receptoru může působit i inhibičně. Převážně je však excitačním mediátorem. Vyskytuje se v korových projekčních neuronech a retikulární formaci. Úbytek se spolupodílí na poruchách paměti (Alzheimerova choroba).
- Monoaminový systém má převážně excitační vliv. S výjimkou adrenergních jsou jejich jádra pigmentovaná. Indolaminy (serotonin, histamin) jsou převážně inhibiční mediátory:
- Adrenergní systém se uplatňuje ve formatio reticulatis.
- Noradrenergní systém je lokalizována ve formatio retikularis a je distribuován do celého CNS. V kůře se podílí na zvyšování pozornosti, ovlivňuje dýchání a kardiovaskulární systém.
- Dopaminergní systém je nutný pro správnou činnost extrapyramidopvého systému. Velké nahromadění je ho v oblasti substantia nigra compacta a je cestou tr. nigrostriaticus dopraovávn do striata. Projevem poruchy z degenerace nigrostriatového systému je Parkinsonova choroba (hypokineticko-hypertonický syndrom – akinesie, tremor, rigor). Při postižení i systému tegmentalis ventralis je přítomna i demence.
- Serotoninergní systém působí v CNS převážně inhibičně (dle typu receptoru). Z refeálních jádrech RF je distribuován do celého CNS. V míše inhibuje neurony sympatiku a přenos bolesti ze zadním rohu, ale aktivuje motoneurony. Ovlivňuje regulaci teploty, sexuální chování, příjem potravy, má vliv na regulaci spánku. Pokles hladiny serotoninu provází endogenní deprese.
Inhibiční aminokyseliny:
Glycin je mediátorem míšních neuronů, jeho blokáda strychninem vyvolává křeče,
Kyselina gama-aminomáselná (GABA) je nejdůležitější inhibiční mediátor, je obsažen v interneruonech a v projekčních neuronech celého CNS.
Excitační aminokyseliny:
Kyselina glutamová a asparagová jsou excitační mediátory pyramidových projekčních neuronů kůry mozku, mozečku, v neuronech a interneuronech sítnice, v hippocampu. Jejich hladiny klesají při Alzheimerově chorobě.
| Obrázky ke kapitole: Spoje centrálního nervového systému | |||
Obr. 71 Mozkové spoje
|
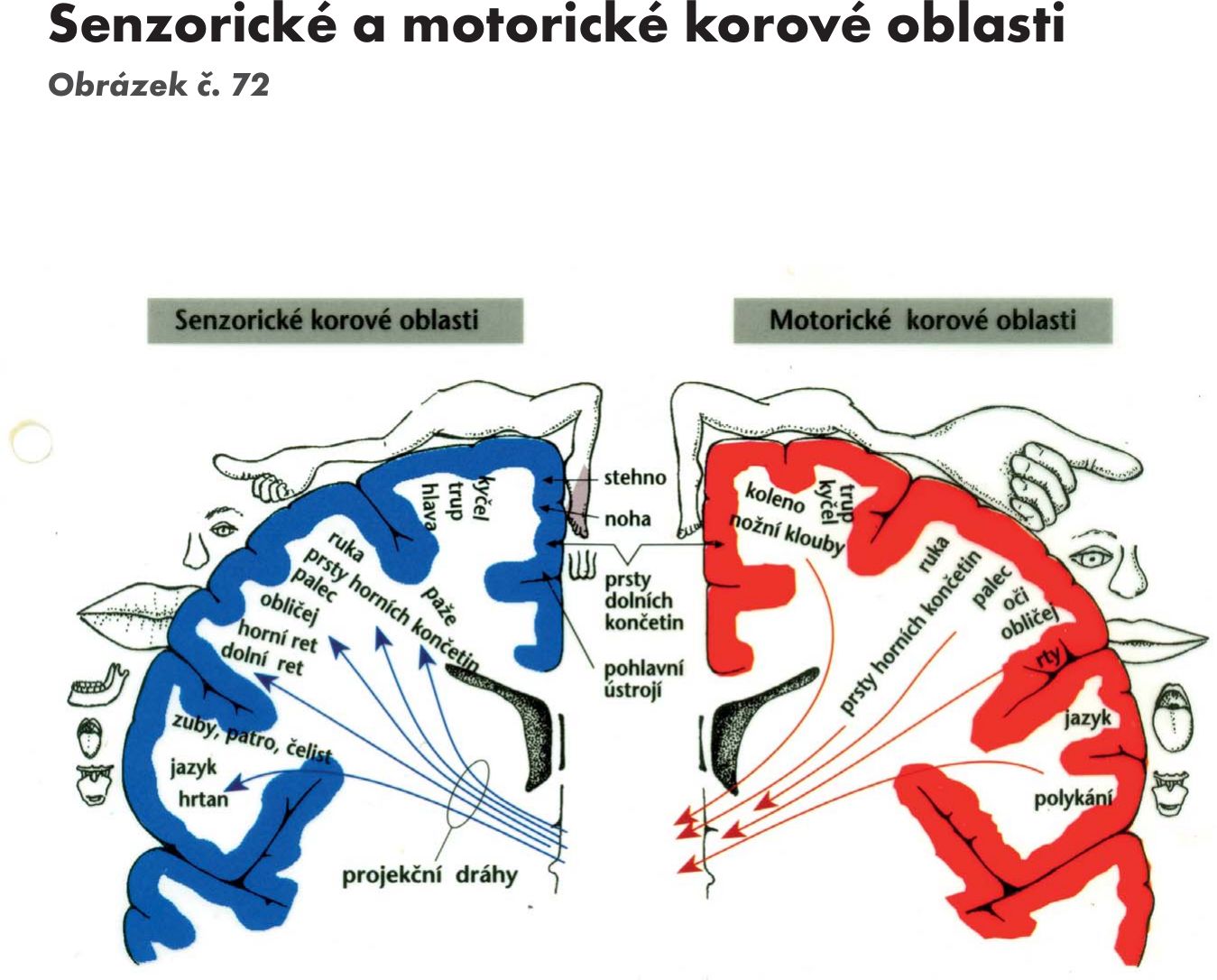
Obr. 72 Senzorické a motorické korové oblasti
|
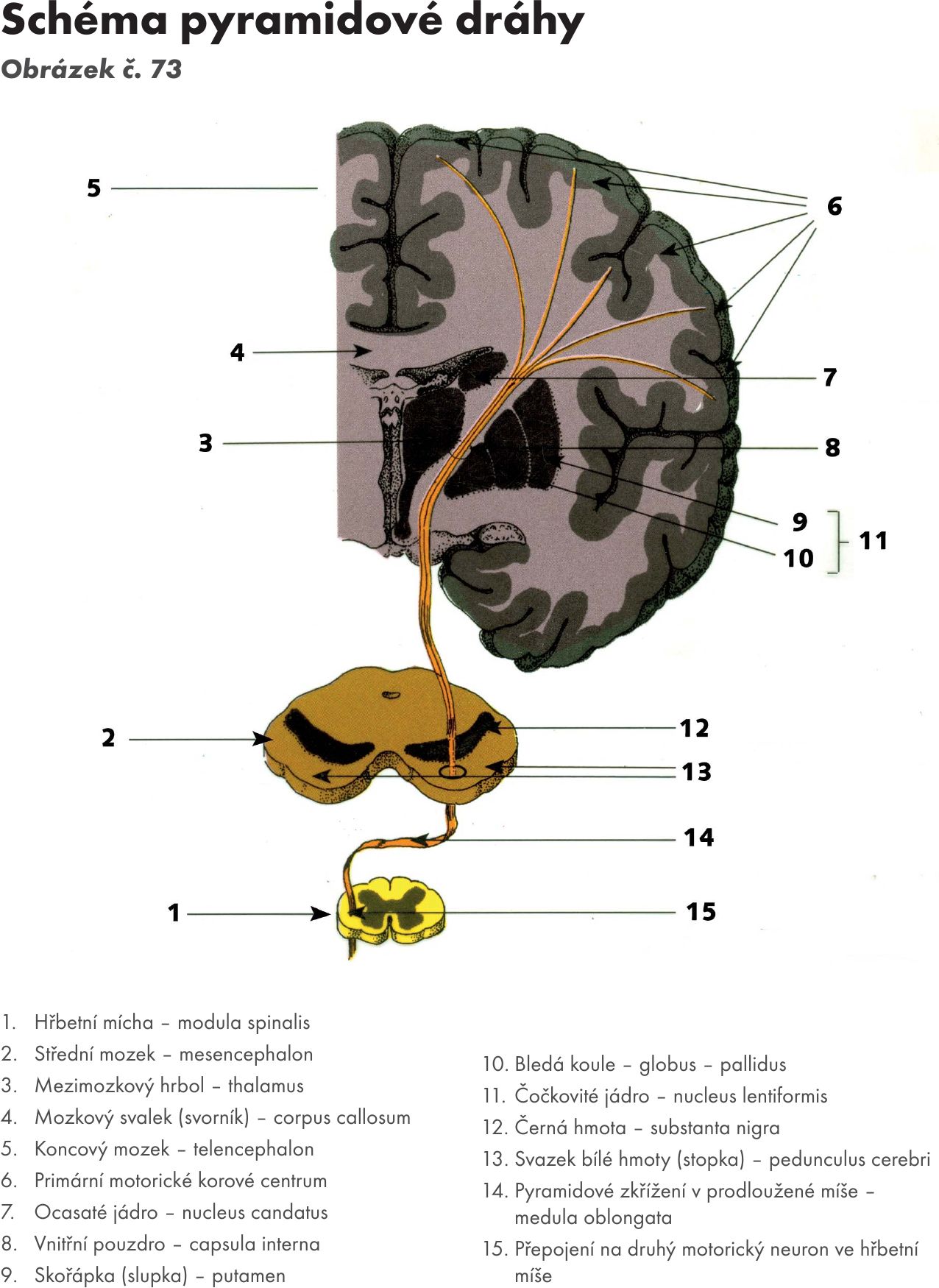
Obr. 73 Schéma pyramidové dráhy
|

Obr. 74 Pyramidové dráhy s připojením na motorických jádrech hlavových nervů
|
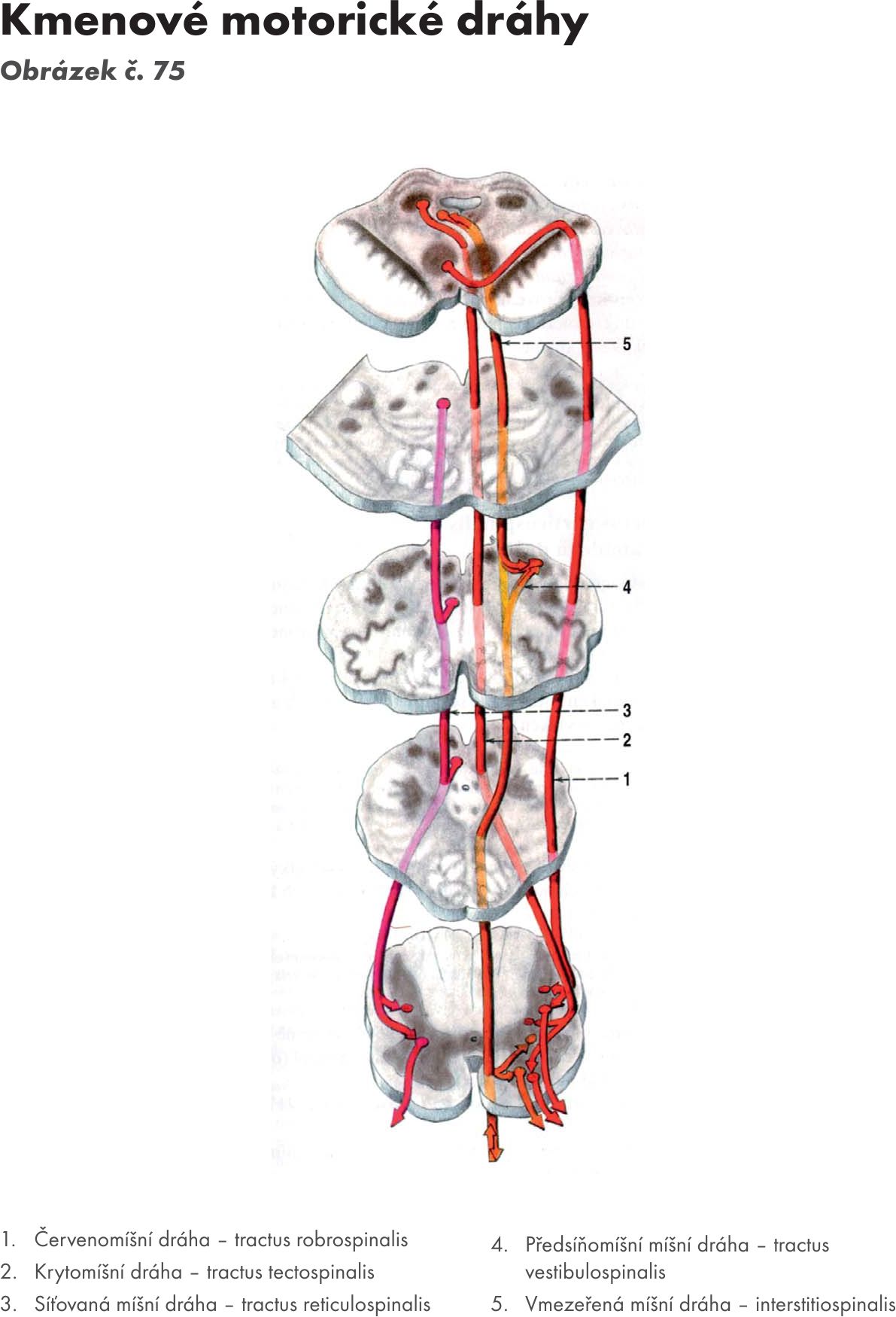
Obr. 75 Kmenové motorické dráhy
|
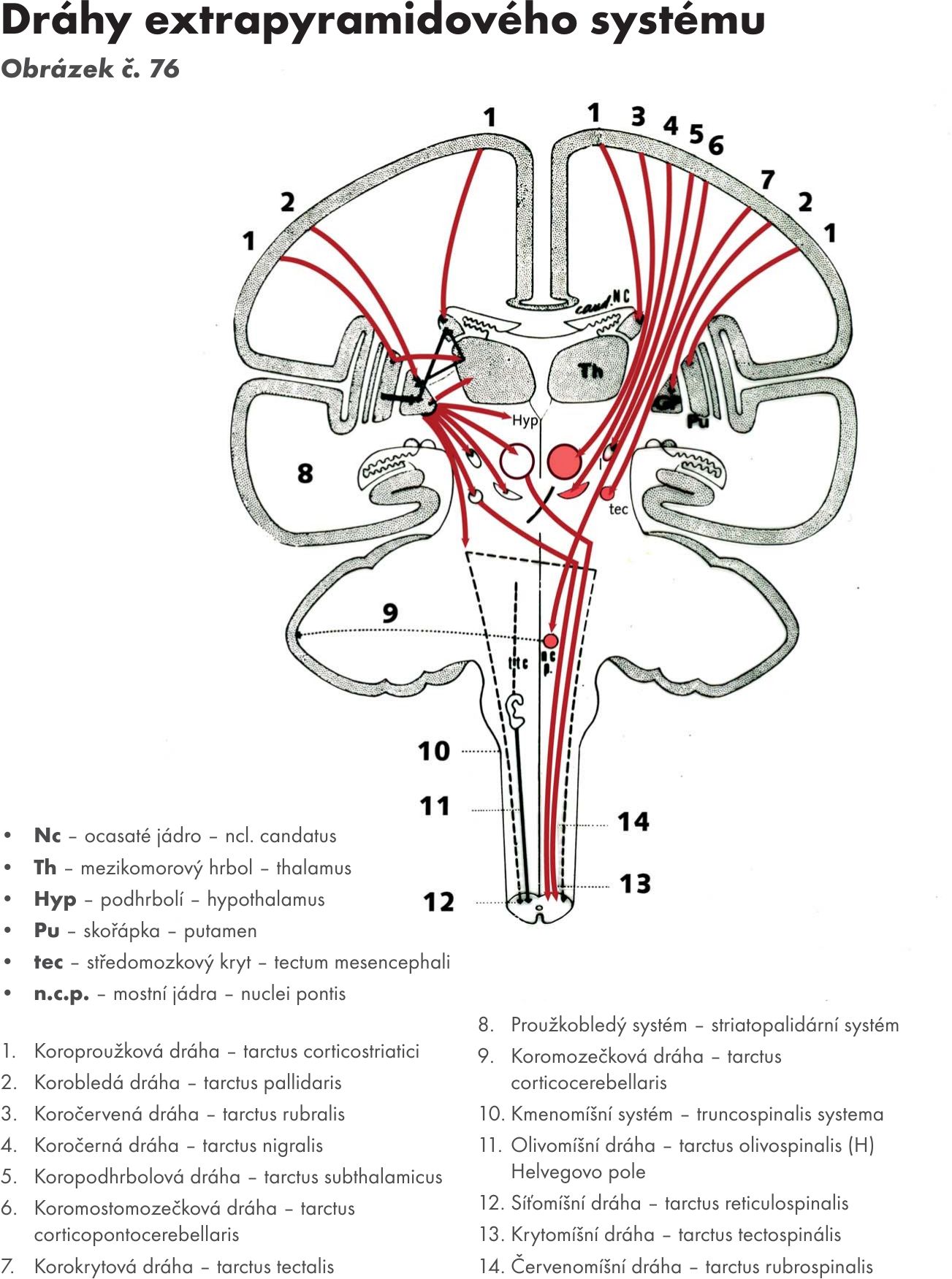
Obr. 76 Dráhy extrapyramidového systému
|
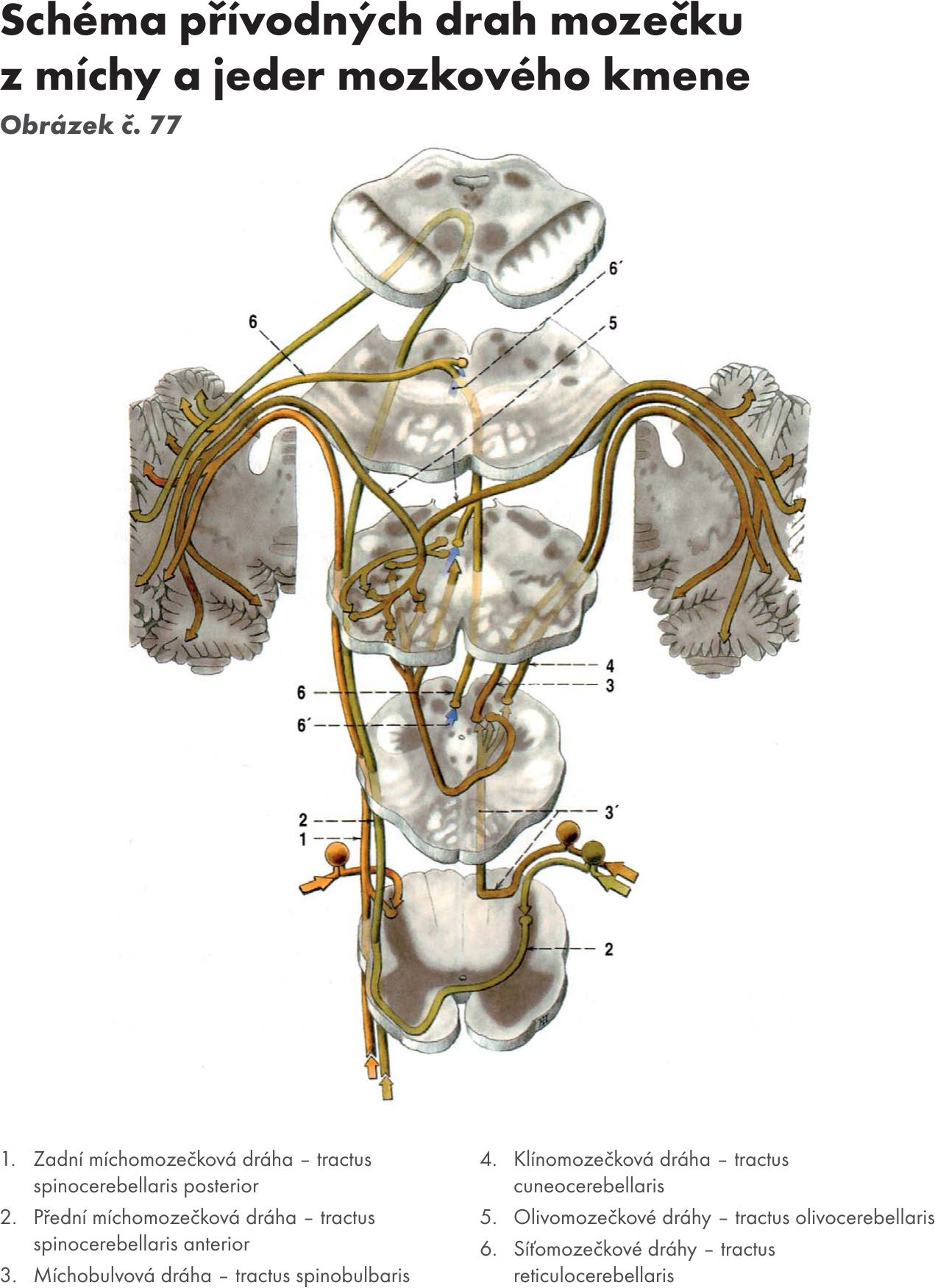
Obr. 77 Schéma přívodných drah mozečku z míchy a jeder mozkového kmene
|

Obr. 78 Přívodné dráhy mozečku z mozkového kmene
|
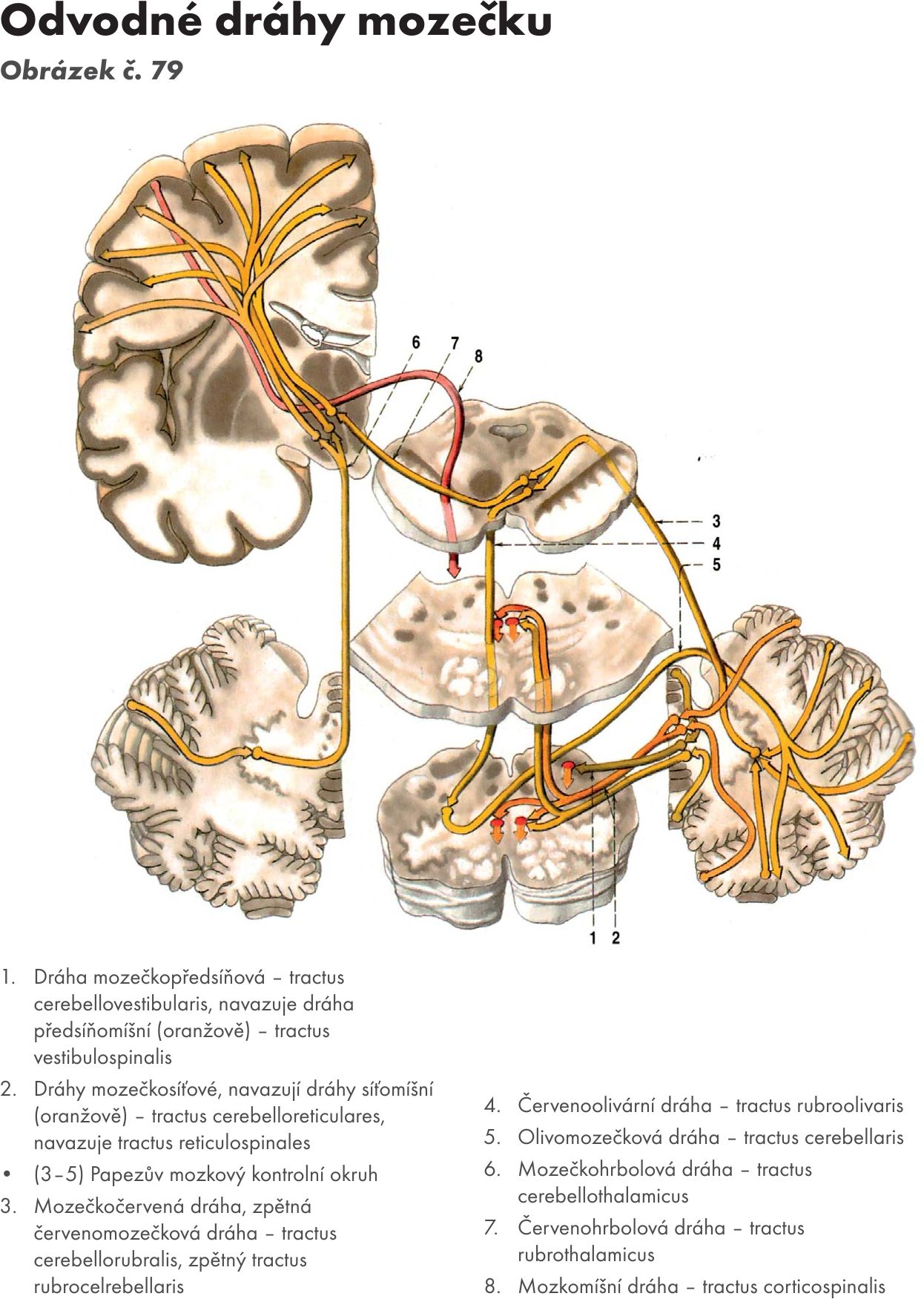
Obr. 79 Odvodné dráhy mozečku
|
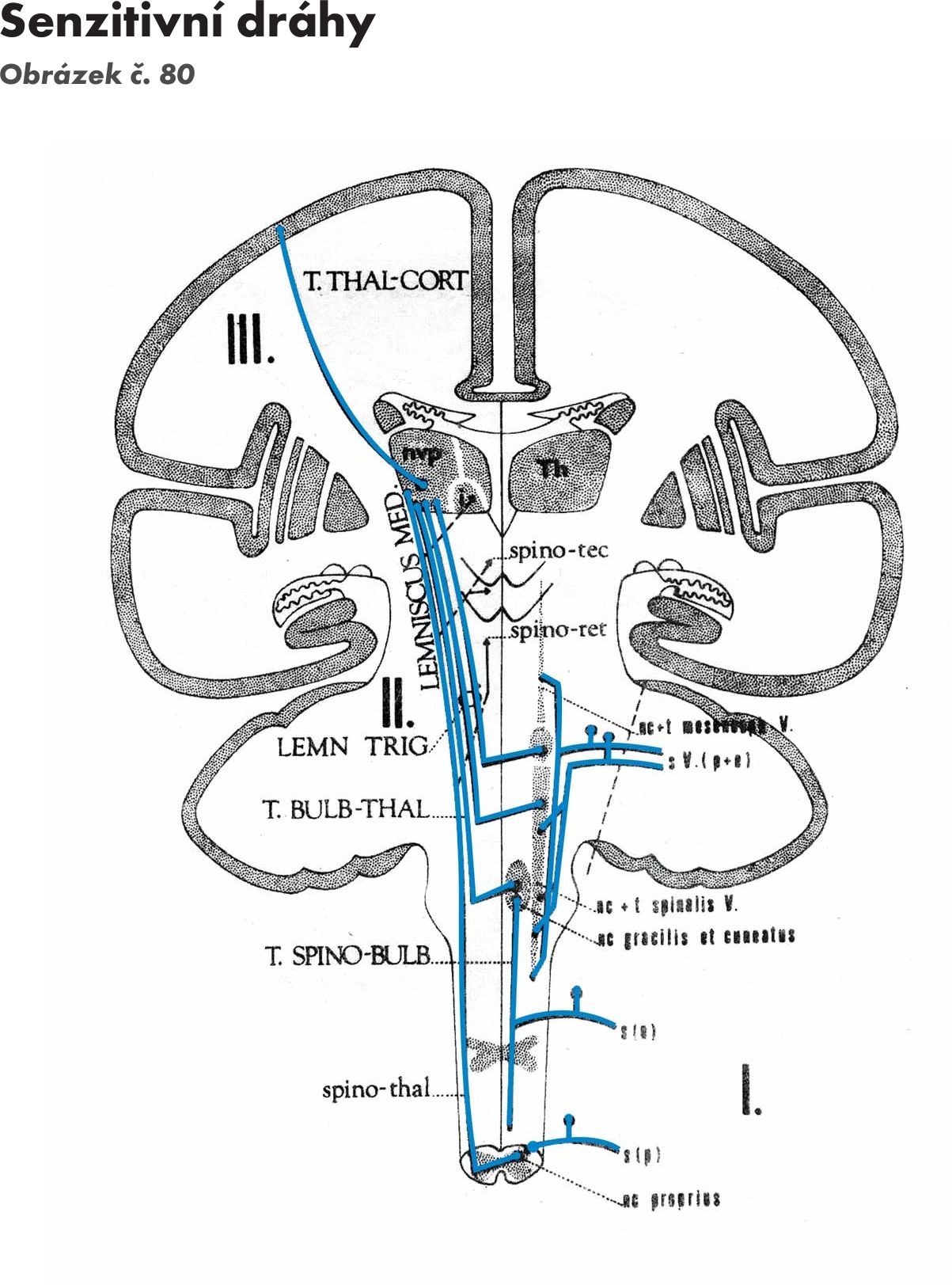
Obr. 80 Senzitivní dráhy
|
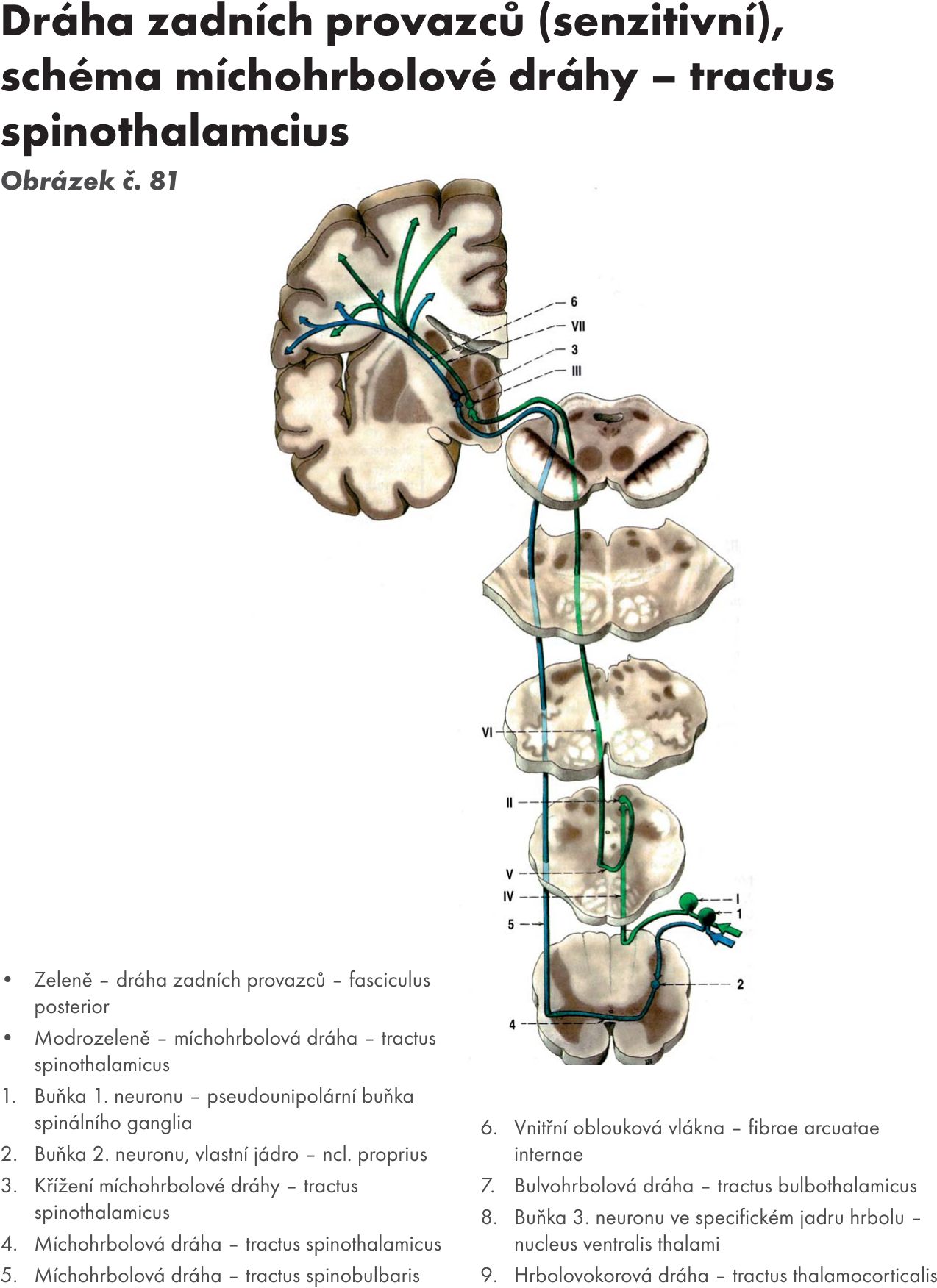
Obr. 81 Dráha zadních provazců (senzitivní), schéma míchohrbolové dráhy – tractus spinothalamcius
|
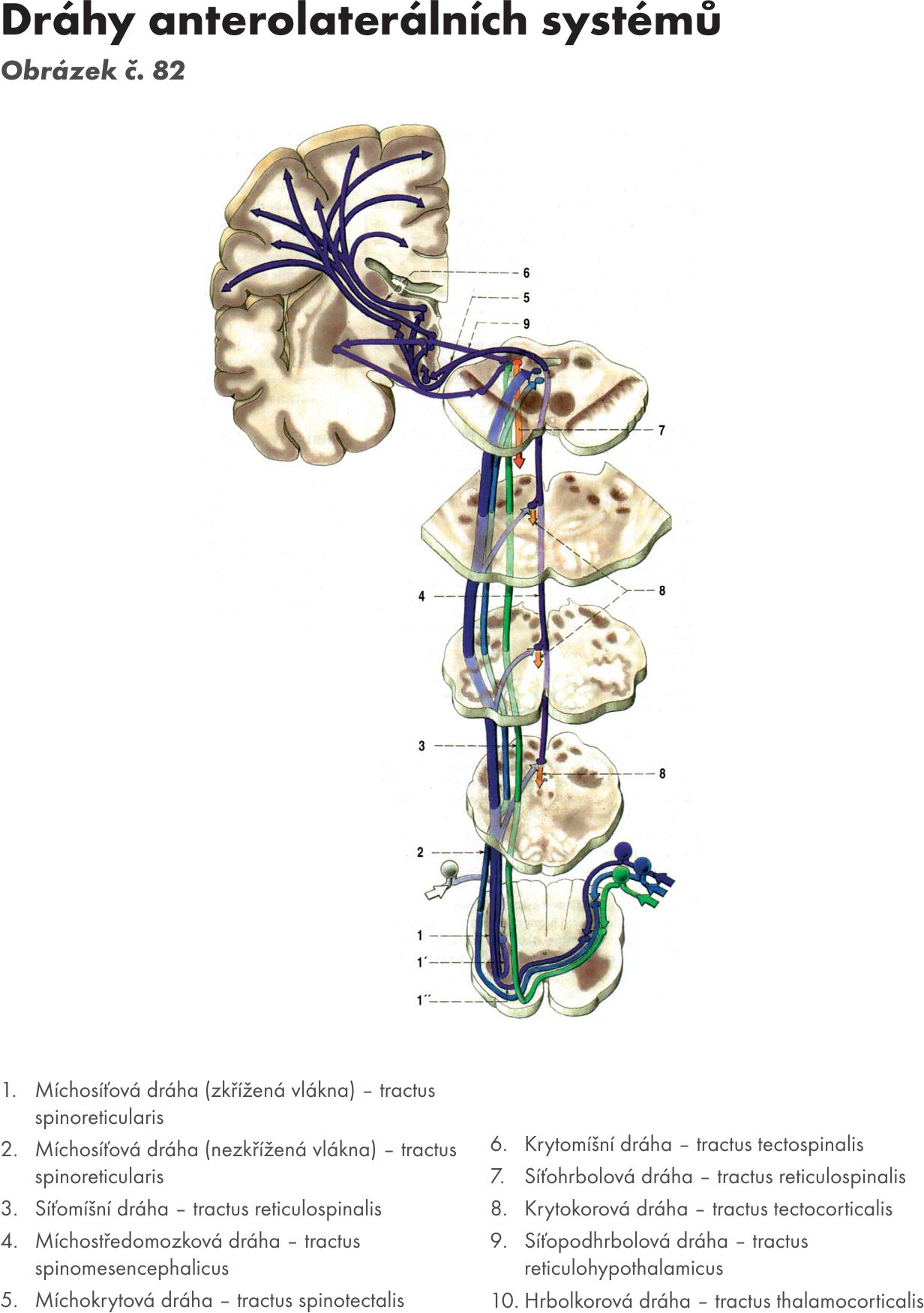
Obr. 82 Dráhy anterolaterálních systémů
|
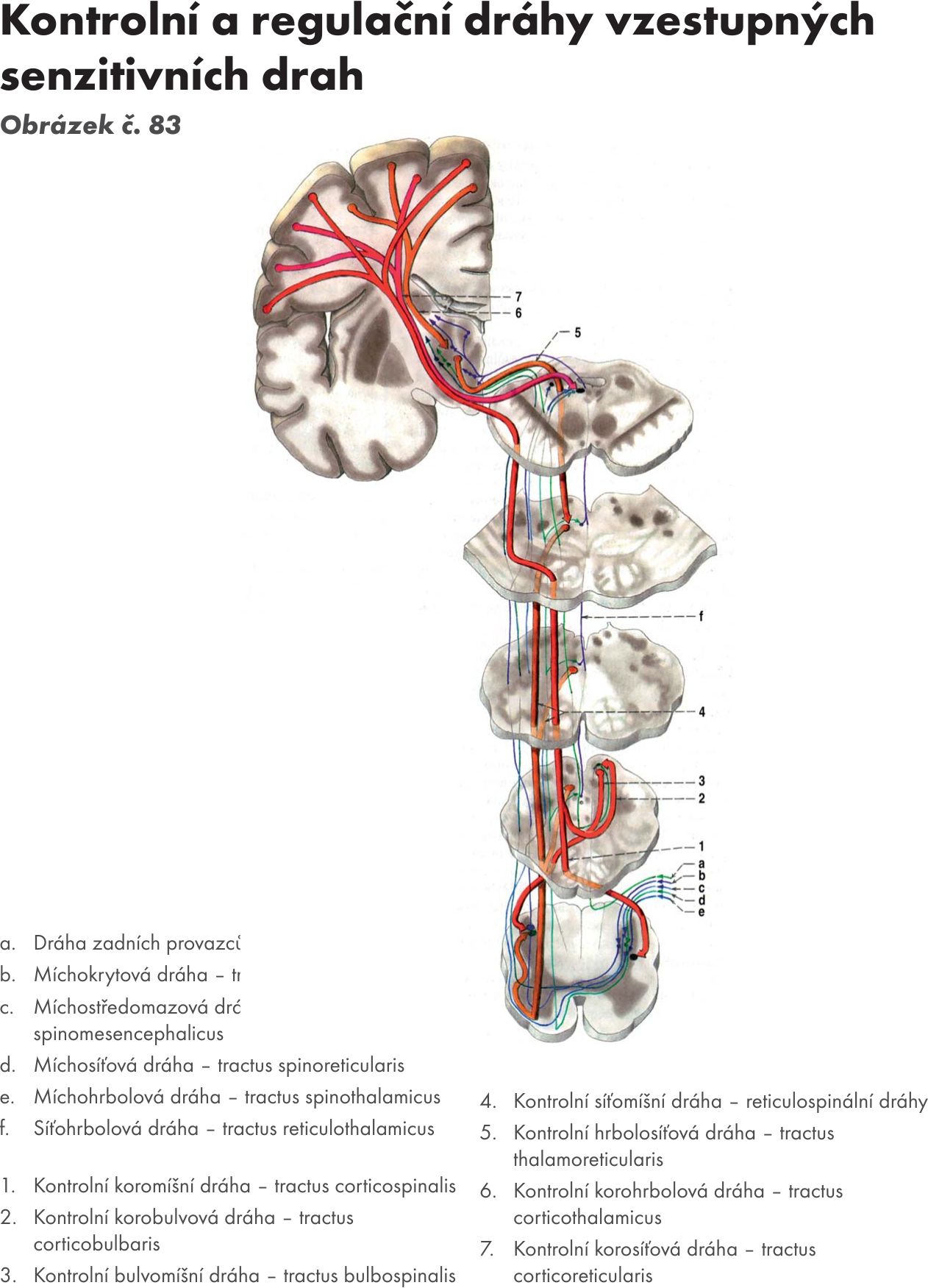
Obr. 83 Kontrolní a regulační dráhy vzestupných senzitivních drah
|
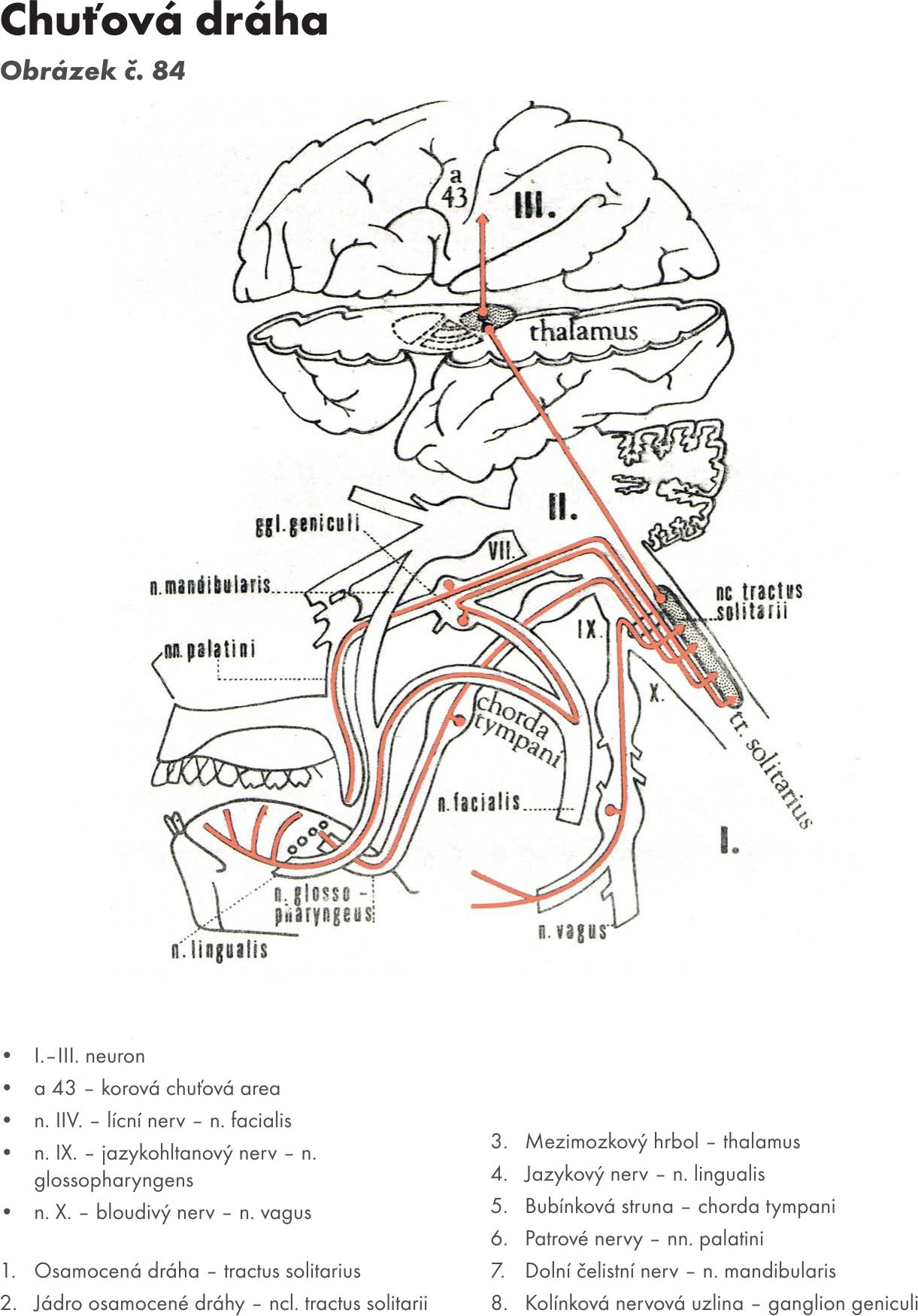
Obr. 84 Chuťová dráha
|
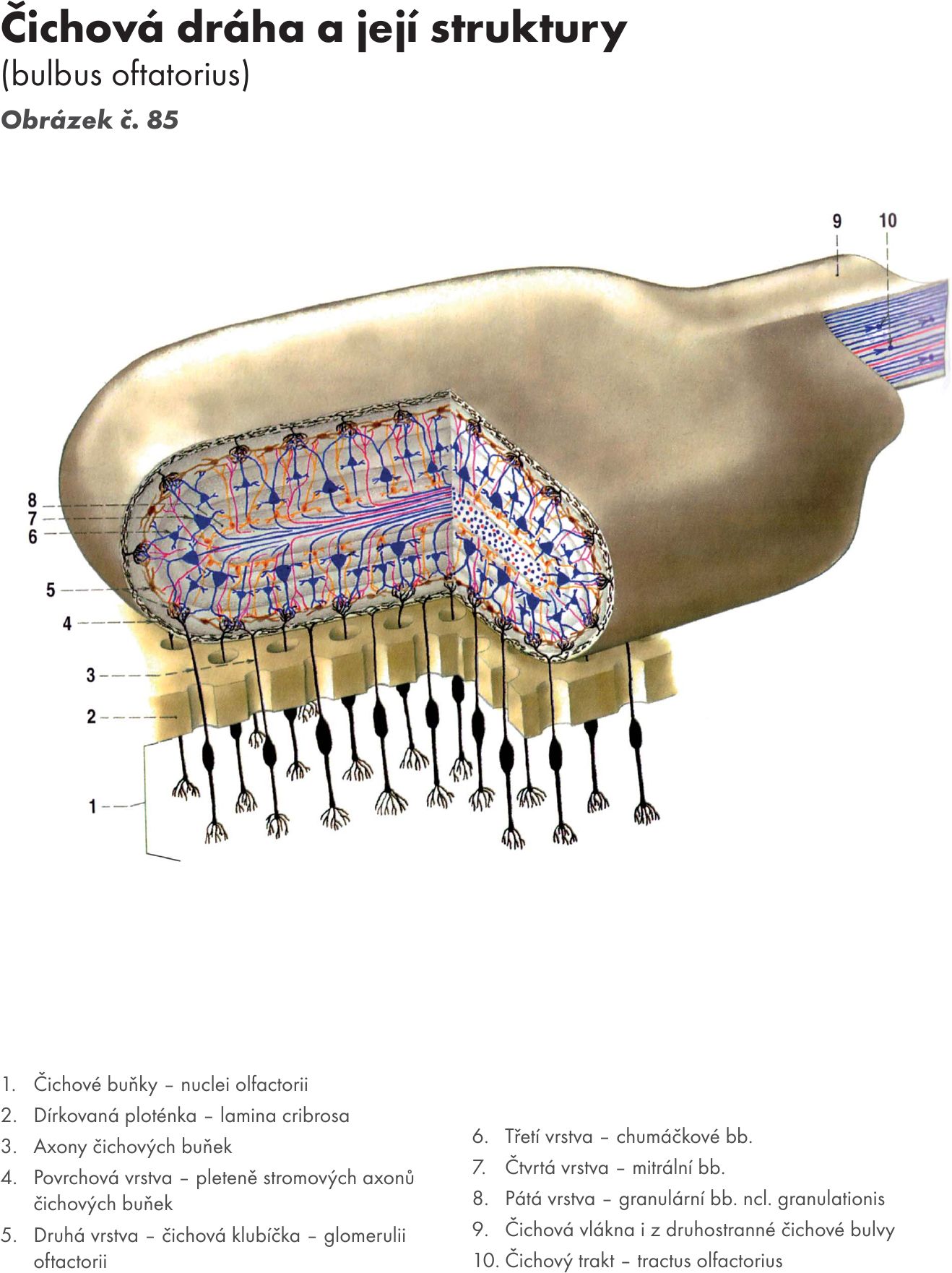
Obr. 85 Čichová dráha a její struktury
|

Obr. 86 Čichová dráha I.-II. neuron
|
KK, Fakulta sportovních studií, Masarykova univerzita |
Návrat na úvodní stránku webu, přístupnost |
| Servisní středisko pro e-learning na MU
| Fakulta informatiky Masarykovy univerzity, 2014
Centrum interaktivních a multimediálních studijních opor pro inovaci výuky a efektivní učení | CZ.1.07/2.2.00/28.0041
