
2.6.2 Sympatrická a parapatrická speciace
Speciace se může vyskytovat také v rámci prostorově se překrývajících populací (bez geografické izolace). K sympatrické speciaci tedy dochází mezi populacemi v jediném areálu. K parapatrické speciaci dochází tehdy, jestliže se v některé populaci z okraje areálu vytvoří odlišnost, která se v dané populaci zafixuje. Díky vzniku reprodukčně izolačních mechanizmů dojde následně k úplnému odštěpení od původního druhu.
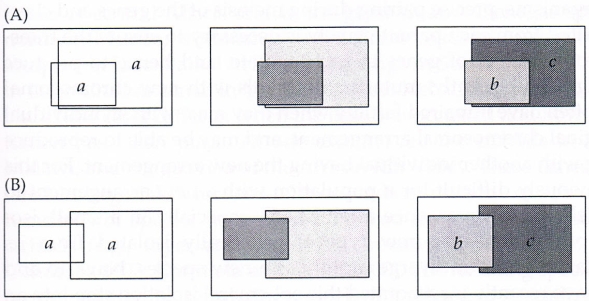
Obr. 29: Ilustrace sympatrické a parapatrické speciace. Při sympatrické speciaci (A) se dvě populace původního druhu a prostorově překrývají a jsou v kontaktu i v průběhu divergence obou populací až do vzniku nových druhů b a c. V případě parapatrické speciace (B) se populace prostorově překrývají pouze v úzké zóně, avšak, stejně jako v předchozím případě, jsou v kontaktu během celého speciačního procesu Brooks & McLennan 2002, Lomolino et al. 2006).
Disruptivní selekce
Pokud silné selekční tlaky způsobí, že se populace adaptují na dva nebo více environmentálních režimů nebo nik, mohou se od sebe postupně oddělit, a nakonec vyústit v nový druh. Disruptivní selekce může být běžná u zvířecích parazitů a v některých skupinách fytofágního hmyzu, které jsou vysoce specializované na konkrétní hostitelské druhy. Mezi těmito organismy je úspěšná kolonizace nového druhu hostitele vzácnou událostí, ale když k ní dojde, kolonisté jsou ihned vystaveni selekci schopnosti přežít a rozmnožovat se v drasticky odlišném prostředí. Obvykle je tento selekční tlak zesílený protikladnou evolucí hostitele, aby unikl nebo odmítl parazita. Zpočátku budou páření mezi hmyzem nebo parazity hostitelsky specifických ras vytvářet potomky, kteří budou mít redukovanou fitness v kterékoli hostitelské nice. Selekce tak bude podporovat vznik prezygotického izolačního mechanismu a dokončení procesu speciace. Podle parapatrického modelu speciace, disruptivní selekce probíhající podél environmentálního gradientu, může postupně přiostřovat klinální variabilitu, dokud nebude původní populace rozdělena na dva nebo více druhů.
Chromozómové změny
K sympatrické nebo parapatrické speciaci může dojít také v důsledku chromosomových změn. Náhodná přestavba genetického materiálu rodiče během meiózy, embrya během oplodnění nebo v raném vývoji, může někdy změnit počet chromozomů nebo pořadí genů na chromosomech. Změny počtu chromozomů jsou dvojího typu: aneuploidie, při které se jeden chromozom rozejde nebo sloučí s jiným. Celkový počet chromozomů tedy bude o jeden větší nebo menší. V případě polyploidie, dojde ke zmnožení celých chromozómových sad, takže v genomu najdeme od každého chromozómu více kopií, např. čtyři, tj. dvě od otce a dvě od matky, vzácně se vyskytují i liché násobky. V ostatních případech zůstává počet chromozomů stejný, ale část genetického materiálu je přestavěna uvnitř chromozomu (inverze) nebo přenesena na jiný chromozom (translokace).
U diploidních organismů mají mutovaní jedinci s novým chromozomovým uspořádáním často oslabenou plodnost, pokud se páří s jedinci, kteří mají původní chromosomové uspořádání. Jsou tedy schopni rozmnožovat se pouze pářením s dalšími jedinci, kteří mají nové uspořádání chromozomů. Proto je těžké založit populaci s novým uspořádáním chromozomů, avšak když je již jednou založena - zejména v malých, izolovaných populacích - nový typ je geneticky izolován od rodičovské populace a může rapidně divergovat v nový druh. Tento proces označujeme jako allo-parapatrický model specice.
Sympatrická speciace v izolovaných jezerech
Jedním z nejpřesvědčivějších důkazů sympatrické speciace u zvířat je divergence a adaptivní radiace ryb v izolovaných jezerech. Dobrým příkladem jsou cichlidy ve Velkých afrických jezerech nebo halančíkovci v Lago Chichancanab na Yucatánu.
Cichlidy v jezerech Malawi, Tanganika a ve Viktoriině jezeře ve Východoafrické příkopové propadlině byly izolovány desítky tisíc až miliony let. Z několika zakládajících vývojových větví cichlid se tak vyvinuly stovky druhů. Výsledné druhy vykazovaly obrovské variace v morfologii, většinou související s různým typem potravy (aby byla snížena kompetice a využity ekologické možnosti); a v barevných vzorech a chování, které souvisejí s projevy dvoření a páření. Poslední důkazy svědčí o tom, že mělké Viktoriino jezero bylo v pozdním pleistocénu kompletně vyschlé. Dokonce, pokud by i jeho úplné vysušení nenastalo, toto jezero pravděpodobně není více než 0,5 až 1 milion let staré a tak fakt, že teď v něm žije více než 300 druhů endemických cichlid, dokládá že při vhodných podmínkách může být speciace a adaptivní radiace extrémně rychlá.
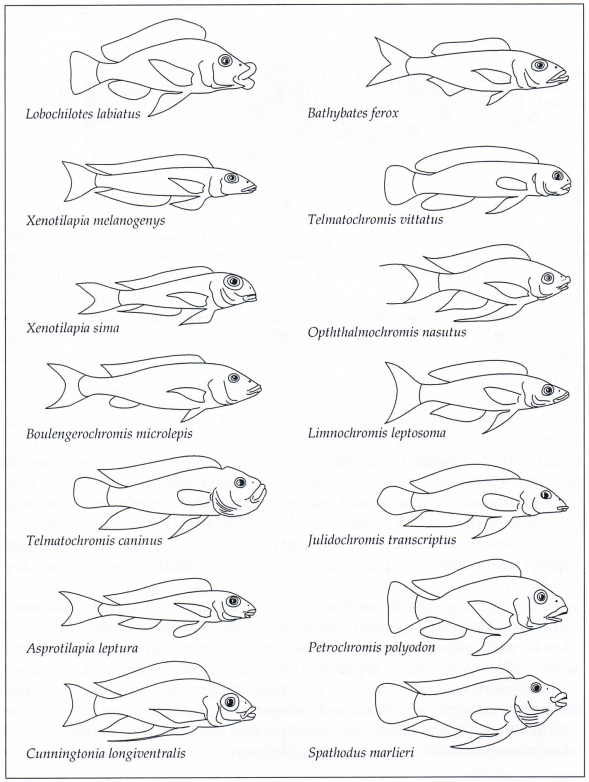
Obr. 30: Příklady rozmanitosti morfologických forem cichlid, které jsou výsledkem procesu adaptivní radiace v jezeře Tanganika ve východní Africe (Fryer & Iles 1972, Lomolino et al. 2006).
Opačným extrémem jsou halančíkovci v Yucatánském Lago Chichancanab, kteří byly izolováni v malých jezerech jen pár tisíc let jako výsledek měnící se hladiny moře a jezer od pleistocénu (izolace slaného jezera od moře). Lago Chichancanab obsahuje pět druhů rodu Cyprinodon (halančíkovec), které mají odlišnou morfologii související se sběrem potravy a také vykazují jistý stupeň genetické a behaviorální reprodukční izolace. Genetické a behaviorální studie ukazují, že proces speciace zde ještě není dokončen a že některé z těchto forem se v určitém rozsahu stále kříží. Rozdíly ve velikosti a tvaru reflektují odlišnosti v potravě a ukazují, že silnou selekci pro trofickou diferenciaci řídí sympatrická speciace.

Obr. 31: Morfologie pěti forem halančíkovců (Cyprinodon) v jezeře Chichancanab na polostrově Yucatán v Mexiku. Tyto formy sympatricky divergovaly během posledních několika málo tisíců let od té doby co bylo jezero izolováno od moře. Genetické a behaviorální studie naznačují, že speciační proces zatím není u konce a že některé tyto formy se stále kříží. Rozdíly ve velikosti a tvaru těla odrážejí rozdílné způsoby obživy a naznačují, že z tohoto hlediska existuje silná selekce, která řídí sympatrickou speciaci (Strecker et al. 1996, Lomolino et al. 2006)
Tyto případy dokazují, že sympatrická
speciace může být běžnější než se většina evolučních biologů
domnívá. Také ukazují, že disruptivní selekce, využití
různých ekologických nik nebo silný pohlavní výběr mohou být
často dostatečně vlivnými silami, aby vyvolaly speciaci,
dokonce i při absenci prostorové izolace. Převládajícím
typem speciace však stále zůstává allopatrická speciace.
<< Předchozí | Nahoru | Následující >>
Zdroje:
BEGON, M., HARPER, J.L. & TOWNSEND, C.R. (1997): Ekologie: jedinci, populace a společenstva. 1. vyd. Vydavatelství Univerzity Palackého, Olomouc. 949 s. ISBN 8070676957
COX, B.C. & MOORE, P. D. eds. (1999): Biogeography: an ecological and evolutionary approach. 6th edition. Oxford: Blackwell Science. ix, 298 s. ISBN 086542778X
BROOKS, R.D. & McLENNAN (2002): The Nature of Diversity: An Evolutionary Voyage of Discovery. University of Chicago Press, Chicago.
FRYER, G. & ILES, T.D. (1972): The Cichlid Fishes of the Great Lakes of Africa: Their Biology and Evolution. Oliver & Boyd. Edinburgh.
KREBS, CH.J. (2001): Ecology: the experimental analysis of distribution and abundance. 5th ed. San Francisco, Calif.: Benjamin Cummings. 695 s. ISBN 0321042891
LOMOLINO, M. V., RIEDLE, B. R., & BROWN, J. H. eds. (2006): Biogeography. 3rd edition. Sunderland, Mass.: Sinauer Associates. xiii, 845. ISBN 0-87893-062-0
MACDONALD, G.M. (2003): Biogeography – Space, Time and Life. John Wiley & Sons, Inc. New York. ISBN 0-471-24193-8
STRECKER, U., MEYER, C.G., STURMBAUER, C. & WILKENS, H. (1996): Genetic divergence and speciation in an extremely young species flock in Mexico formed by the genus Cyprinodon (Cyprinodontidae, Teleostei). Molecular Phylogenetics and Evolution 6: 143-149.
- Autoři:
- Mgr. Jan Divíšek
- RNDr. Martin Culek, Ph.D.
- Mgr. Martin Jiroušek
- © 2010
- Geografický ústav, Přírodovědecká fakulta, Masarykova univerzita
- Tvorba této příručky byla podpořena projektem FRVŠ 2209/2010/G4 Zkvalitnění výuky předmětu Z0005 Biogeografie
- Návrat na úvodní stránku, Nahoru
- El-portál