
2.5.5 Adaptace a tok genů
Nika druhu není konstantní v prostoru ani v čase. Stejně jako je přírodní výběr univerzální proces, který má tendenci zvyšovat schopnost jedinců na přežití a rozmnožování, čekali bychom, že populace se adaptují na svoje místní prostředí. Taková adaptace se může projevovat geografickou variabilitou uvnitř druhu ve fyziologii či morfologii. Můžeme očekávat, že periferní populace budou schopné adaptovat se na environmentální faktory, které limitují jejich rozsah, zvýší se jejich hustota a jedinci kolonizují přilehlé oblasti.
Během posledního století queenslandská ovocná muška – Bactrocera (Dacus) tryoni rozšířila svůj areál o několik stovek kilometrů na jih podél východního okraje kontinentu. Tyto mušky jsou limitovány nízkými teplotami a jejich expanze byla doprovázená adaptací periferní populace na vzrůstající toleranci k chladu.
Proč se tedy na lokální podmínky neadaptují periferní populace všech druhů s postupnou expanzí rozsahu do všech periferií? V některých případech je důvod zřejmý. Existují zásadní omezení, která nemohou být překonaná lokální adaptací. Například pevninský život, který se formuje podél pobřeží, si nemůže jednoduše vyvinout adaptaci na vodní život a proniknout do mořské říše. Tento druh adaptace se sice uskutečnil v minulosti – když suchozemští předkové tučňáků a velryb pronikli do moře – ale trvalo to miliony let a vyžadovalo to, aby se tato zvířata vzdala své schopnosti žít na pevnině.
Genetické, ekologické a biogeografické procesy se vzájemně ovlivňují, aby limitovaly schopnost periferních populací adaptovat se na lokální podmínky. Výměna genů mezi populacemi přes disperzi a tok genů může zabraňovat lokálním populacím nabízet a sdílet kombinace genů, nevyhnutelné pro ustavičnou adaptaci. Dostatečně vysoký tok genů může zaplavit lokální okrajovou populaci geny z "mateřské" popuplace, efektivně pracujícími proti vzrůstající adaptaci na lokální podmínky.
Příkladem adaptivní genetické změny v odezvě na variabilitu prostředí je křeček dlouhoocasý (Peromyscus maniculatus), který obývá většinu severoamerického kontinentu. Vyskytuje se tu mnoho variací v barvě a tvaru těla křečka, které se odrážejí v rozdělení druhu do 50 geografických ras či poddruhů. Variace jsou adaptacemi na lokální podmínky a každá rasa má odlišné charakteristiky. Barva srsti se shoduje s lokální barvou půdy, kvůli tomu, že predátoři si vybírají kontrastně zbarvené jedince. V porovnání s pouštními jedinci a jedinci z travnatých stepí mají zvířata žijící v lesních oblastech delší zadní nohy a ocasy, které používají na lezení.
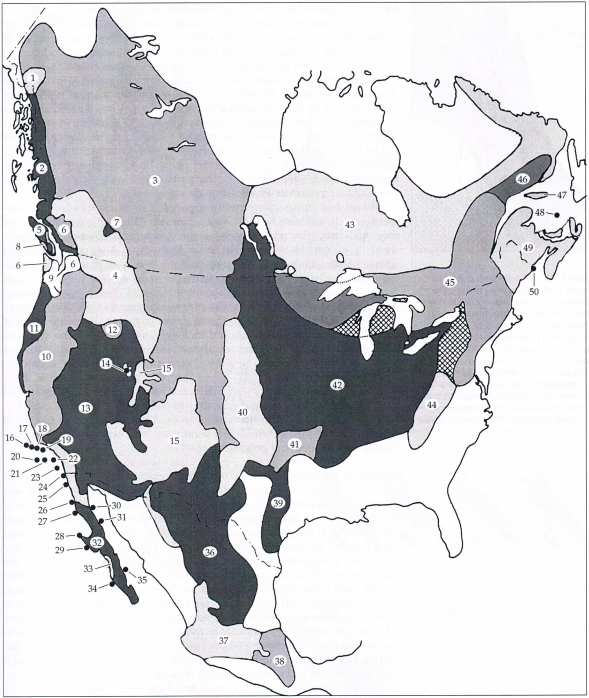
Obr. 27: Geografické rasy křečka dlouhoocasého (Peromyscus maniculatus) jsou známkou rozdělení druhu do 50 formálně uznaných poddruhů. Každá z těchto geografických ras nese zřetelně odlišné znaky zahrnující barvu hřbetu (odpovídá barvě půdy), délku ocasu a zadních noh (odráží typ habitatu a tedy nutnost šplhat)(Hall 1981, Lomolino et al. 2006).
Další příklad adaptace podal, pomocí experimentů v běžné zahradě, Clausen et al. (1940, 1947, 1948), který demonstroval společný vliv genetické izolace, lokální selekce a toku genů na adaptivní odlišnosti lokálních populací řebříčku obecného (Achillea millefolium). Tyto adaptace dovolují řebříčku mít široké rozšíření podél výškového gradientu v horách Sierra Nevady v Kalifornii. Poté, co rostliny z různých nadmořských výšek vyrostly identických podmínkách, bylo možné se domnívat, že rozdíly mezi nimi jsou genetického původu. Podle všeho tyto odlišnosti reflektují lokální adaptace, vzniklé kombinaci přírodního výběru a redukovaného toku genů. Tok genů může bránit lokálním populacím adaptovat se na odlišné lokální podmínky, ale pokud je tento tok přerušen (populace jsou izolovány), potom se v průběhu evoluce mohou populace často lišit v odpovědi na přírodní výběr, adaptovat se na široce odlišné podmínky, expandovat na nová stanoviště a rozšiřovat svůj areál.

Obr. 28: Morfologická diferenciace řebříčku obecného (Achillea millefolium) podél gradientu nadmořské výšky horách Sierra Nevada v Kalifornii. Od té doby co byly tyto rostliny pěstovány v identických podmínkách běžné zahrady se můžeme domnívat, že rozdíly mezi nimi jsou genetického původu. Tyto odlišnosti pravděpodobně reflektují lokální adaptace vzniklé vlivem přírodního výběru a redukovaného toku genů (Clausen 1948, Lomolino et al. 2006).
<< Předchozí | Nahoru | Následující >>
Zdroje:
BEGON, M., HARPER, J.L. & TOWNSEND, C.R. (1997): Ekologie: jedinci, populace a společenstva. 1. vyd. Vydavatelství Univerzity Palackého, Olomouc. 949 s. ISBN 8070676957
CLAUSEN, J., KECK, D.D. & HIESEY, W.M. (1940): Experimental Studies of the Nature of Species. I. Effect of Varied Environments on Western North American Plants. Carnegie Institute Publication 520. Carnegie Institute, Washington DC.
CLAUSEN, J., KECK, D.D. & HIESEY, W.M. (1947): Heredity of geographically and ecologically isolated races. American Naturalist 81: 114-133.
CLAUSEN, J., KECK, D.D. & HIESEY, W.M. (1948): Experimental Studies of the Nature of Species. III: Environmental Responses of Climatic Races of Achillea. Carnegie Institute Publication 5581. Carnegie Institute, Washington DC.
COX, B.C. & MOORE, P. D. eds. (1999): Biogeography: an ecological and evolutionary approach. 6th edition. Oxford: Blackwell Science. ix, 298 s. ISBN 086542778X
HALL, E.R. (1981): The Mammals of North America. 2nd edition. 2 vols. John Wiley & Sons, New York.
KREBS, CH.J. (2001): Ecology: the experimental analysis of distribution and abundance. 5th ed. San Francisco, Calif.: Benjamin Cummings. 695 s. ISBN 0321042891
LOMOLINO, M. V., RIEDLE, B. R., & BROWN, J. H. eds. (2006): Biogeography. 3rd edition. Sunderland, Mass.: Sinauer Associates. xiii, 845. ISBN 0-87893-062-0
MACDONALD, G.M. (2003): Biogeography – Space, Time and Life. John Wiley & Sons, Inc. New York. ISBN 0-471-24193-8
