
4.3 Růst populace
Skutečná míra disperze druhu do nového území je ovlivněna jak vzdáleností, tak i povahou překážek a samozřejmě i vlastnostmi samotného šířícího se druhu. Téměř pro všechny organismy má míra imigrace (počty jedinců nebo druhů, kteří přibydou na určitém místě za jednotku času) tendenci klesat s rostoucí vzdáleností od zdroje migrantů. Pokud má mít šíření druhu význam, jedinci druhu musí být schopni se na nové stanoviště nejen dostat, ale hlavně zde vytvořit životaschopnou populaci. Dostat se na novou lokalitu je proto jenom poloviční výhra, neboť úspěšný kolonista je ten, kdo přežívá a rozmnožuje se. Kolonizace je tedy sled událostí, zahrnující nejen disperzi propagule do nového území, které ještě nebylo daným druhem osídleno, ale také úspěšné založení reprodukceschopné populace.
U organismu, který kolonizuje novou oblast a neomezeně využívá místní zdroje, předpokládáme exponenciální růst jeho populace. Například sameček a samička pěnkavy se dostanou na nové lokality na velkém ostrově, kde se rozmnožují. Další rok rodičovský pár zplodí dva samečky a dvě samičky. Na ostrově teď žijí 3 páry pěnkav, které zplodí celkově dalších 12 potomků. V poměrně krátkém časovém intervalu činí celková populace 18 jedinců. Tento proces se nazývá exponenciální růst populace a může být popsán vzorcem
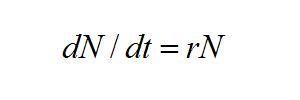
kde dN je změna velikosti populace (N), dt se rovná časovému přírůstku (v tomto případě jedna generace) a r je míra populačního růstu na hlavu (počet potomků každého rodiče mínus počet jedinců zemřelých před reprodukcí).
Zdroje na ostrově, který pěnkavy osídlily, však ve skutečnosti nejsou neomezené. Jak se velikost populace postupně přibližuje únosné kapacitě prostředí, zvyšuje se vnitrodruhová kompetice o zdroje a dochází tak ke zpomalení míry populačního růstu. Pokud jsou brány v úvahu konečné zdroje, populační růst kolonizujícího druhu má tvar logistické křivky. Logistický růst populace lze matematicky popsat jako
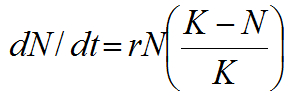
kde K je únosná kapacita prostředí. Jakmile se N blíží K, míra populačního růstu se zpomalí. Když N=K je populační růst nulový, neboť únosná kapacita prostředí je naplněna. Logistický model lépe přibližuje skutečný růst populace kolonizujících druhů, protože na rozdíl od exponenciálního modelu uvažuje omezené zdroje.

Obr. 66: (A) Křivky ilustrující exponenciální a logistický růst populace. (B) Příklad logistického růstu populace, který byl pozorován v laboratorních podmínkách u korovníka obilného (Rhyzopertha dominica). (C) Příklad logistického růstu populace u různých druhů smrku (Picea), které kolonizovaly oblast západní Kanady během holocénu. Populační růst je odhadnut na základě palynologických dat z jezerních sedimentů (Crombie 1945, MacDonald 1993, MacDonald 2003).
Dalším způsobem porovnání populačního růstu různých populací je výpočet doby zdvojení populace. Je to počet let, který je potřeba ke zdvojnásobení velikosti populace:
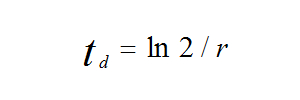
kde td je doba potřebná ke zdvojnásobení populace, r je míra populačního růstu na hlavu a ln2 (přirozený logaritmus dvou) se rovná přibližně 0,70.
Logistická rovnice populačního růstu kolonizujících druhů byla testována laboratorními experimenty na jednobuněčných organismech, jako jsou kvasinky, a dále také na hmyzu, jako např. na octomilce obecné (Drosophila melanogaster), hrotnatce velké (Daphnia magna), broucích (Tribolium a Rhizopertha) nebo na jednoduchých rostlinách (Spirodela oligorrhiza). Logistický růst populace kolonizujícího druhu však podporují také skutečná pozorování mimo laboratorní podmínky. Například studium primární sukcese na vulkanickém ostrově Surtsey nedaleko Islandu ukázalo, že kolonizace holých skal mechem sleduje jasný logistický trend. Pylové analýzy rovněž ukazují logistický růst populace některých druhů stromů osídlujících odledněnou krajinu v Severní Americe a Asii na začátku holocénu.
Kromě hladkého esovitého logistického růstu může dojít k případům, kdy populační růst pokračuje přes průměrnou únosnou kapacitu prostředí. Populace pak následně zažívá katastrofický pokles k hranici únosné kapacity nebo ještě níže. Tento jev se nazývá překročení únosné kapacity prostředí. Tento jev lze dokumentovat na příkladu populace soba polárního (Rangifer tarandus). V roce 1911 byli na ostrov Sv. Pavla v Beringově moři introdukováni 4 samci a 21 samic kvůli chovu na maso, kůže a na export. Počáteční populační růst sobů byl exponenciální a do roku 1938 zde bylo více než 2000 sobů. Poté však nastal prudký pokles v populačním růstu, který byl následován poklesem celé populace, a již v r. 1950 celková populace sobů dosáhla počtu pouhých 8 sobů. Tento prudký pokles byl pravděpodobně způsoben nedostatkem potravy, protože zpočátku strmý nárůst počtu sobů způsobil narušení celého ekosystému ostrova, došlo k devastaci porostů. Často také dojde k rozšíření chorob, které prudce narostlou populaci zdecimují.
<< Předchozí | Nahoru | Následující >>
Zdroje:
CROMBIE, A. C. (1945): On competition between different species of garminivorous insects. Proceedings of the Royal Society of London, Series B 132, 362-395.
LOMOLINO, M. V., RIEDLE, B. R., & BROWN, J. H. eds. (2006): Biogeography. 3rd edition. Sunderland, Mass.: Sinauer Associates. xiii, 845. ISBN 0-87893-062-0.
MACDONALD, G. M. (1993): Reconstructing plant invasions using fossil pollen analysis. Advances in Ecological Research 24, 67-110.
MACDONALD, G. M. (2003): Biogeography – Space, Time and Life. John Wiley & Sons, Inc. New York. ISBN 0-471-24193-8
