
2.6 Typy speciace
2.6.1 Allopatrická (geografická) speciace
Geografická izolace je považována za nejčastější, pokud ne nezbytný, první krok v procesu speciace, protože je to nejjednodušší způsob, jak přerušit tok genů mezi populacemi, a nakonec vede k formaci reprodukčních bariér.
Pokud je prostředí heterogenní, geograficky široce rozšířená populace bude mít tendenci vyvinout regionální genetické rozdíly v odezvě buď na přírodní výběr, nebo na genetický drift. Protože však existují bariéry, které limitují disperzi, volný tok genů se vyskytuje jen zřídka. Populace žijící v environmentálně odlišných a geograficky oddělených oblastech tak mají tendenci odlišit se od ostatních, nicméně tok genů zachovává genetickou soudržnost druhu. Příkladem takové diferenciace jsou regionální rasy domácích vrabců, křečka dlouhoocasého (Peromyscus maniculatus) nebo též klinální variace ve velikosti vrhu a snůšky.
Pokud se však regionální populace stanou dostatečně izolovanými, soudržný tok genů mezi nimi je přerušen nebo drasticky redukován, stanou se nezávislými evolučními jednotkami. Například, Darwin poukazoval na morfologické rozdíly mezi obřími želvami na Galapágách, které zjevně pocházejí ze společného předka, ale v současnosti se vyskytují na různých ostrovech.
Allopatrická speciace způsob I: vikariance
Environmentální změna může vytvořit bariéru uvnitř areálu a izoluje tak dříve spojené a vzájemně se křížící populace (např. stoupající hladina moří, tektonické události apod.). Obvykle jsou izolovány relativně velké skupiny populací.

Obr. 30: Ilustrace procesu allopatrické speciace - vikariance - která ústí ve fylogenetický vzor. (A) Původní druh a je široce rozšířený přes celé území. Zformováním geografické bariéry dojde k rozdělení populací tohoto druhu - vikariance. (B) V důsledku přerušeného toku genů, oddělené populace postupně divergují a tato divergence může časem vyústit ve dva odlišné druhy b a c. (C) Jiná geografická bariéra následně rozdělí populace druhu c a ty pak dále divergují až ke vzniku nových druhů d a e (Brooks & McLennan 2002, Lomolino et al 2006).
Gondwanská vikariance
Biogeografové dlouho uvažují, že rozpad Gondwany vedl ke speciaci způsobené vikariancí původního taxonu, který mohl být rozšířen na dvou nebo více fragmentech původního superkontinetu. Příkladem je molekulární vývoj podčeledi žab v rámci čeledi Ranidae. Podčeleď Mantellinae je zcela endemická na Madagaskaru, zatímco její sesterská podčeleď Rhacophorinae má největší rozmanitost v orientální oblasti a nejpravděpodobněji byla součástí bioty před připojením Indie k Eurasii začátkem kenozoika. Oddělení podčeledi Mantellinae nastalo cca před 88 mil. let, kdy došlo ke geologickému oddělení Madagaskaru od pevniny Indie-Seychely.
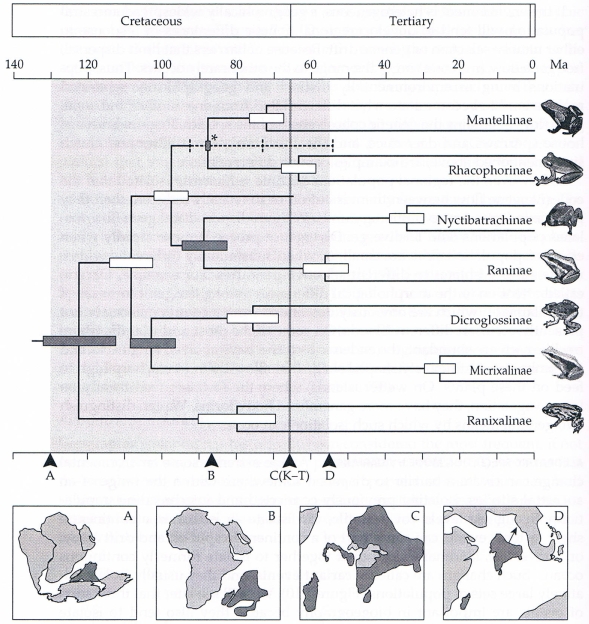
Obr. 31: Molekulární fylogeneze žabích podčeledí náležejících do čeledi skokanovitých (Ranidae). Uzel s hvězdičkou (*) reprezentuje dobře datované geologické oddělení Madagaskaru od kontinentu Indie-Seyschely před 88 mil. let. Tento uzel také ukazuje oddělení Mantellinae - endemické podčeledi Madagaskaru - a její sesterské podčeledi Rhacophorinae, které byly pravděpodobně součástí původní Indicko-Seychelské bioty. Shoda mezi časem oddělení evolučních linií a časem rozdělení pevninských celků nám poskytuje dobrý příklad speciace, která byla vyvolána vikariantní událostí. Mapky (A) až (D) ukazují uspořádání několika Gondwanských pevninských celků od doby oddělení Madagaskaru a celku Indie-Seychely před 130 mil. let (A) až po kolizi Indie s Eurasií před 56 mil. let (D) (Bossuyt & Milinkovitch 2001, Lomolino et al. 2006).
Allopatrická speciace způsob II: periferní izoláty nebo efekt zakladatele
Jedinci se mohou rozšířit přes stávající bariéru, aby kolonizovali předtím neosídlený region. Takové příležitostné a nahodilé události se nazývají "skokové šíření" (jump dispersal) nebo efekt zakladatele (founder effect). Tímto způsobem byly kolonizovány oceánické ostrovy a mnoho jiných izolovaných stanovišť. Typicky se to týká malých počátečních populací, někdy jen jednoho nebo páru jedinců. Na rozdíl od vikariance se tyto události většinou netýkají většího množství druhů zároveň. Jak bylo naznačeno výše, mechanismus a míra počáteční genetické divergence se může lišit v závislosti na způsobu izolace. Genetický drift zde může hrát větší roli než selekce a divergence může být rychlejší, přinejmenším zpočátku, než při vikariaci.

Obr. 32: Ilustrace allopatrické speciace druhého typu a výsledný fylogenetický vzor. (A) Náhodné šíření jedinců široce rozšířeného druhu a do okrajových oblastí, kde zakládají izolované populace. V průběhu času diverguje každá nová periferní izolovaná populace v nový druh b, c a d. (B) Periferní izoláty jsou v tomto případě zformovány postupně zakladateli. Z první izolované populace vznikne druh b, poté někteří jedinci druhu b založí novou izolovanou populaci, která vede ke vzniku druhu c. Následně několik jedinců druhu c založí další izolovanou populaci, ze které postupem času vznikne druh d (Brooks & McLennan 2002, Lomolino et al. 2006).
Allopatrická speciace na souostroví Galapágy
Mezi populacemi želv na každém z velkých Galapážských ostrovů jsou zřetelné morfologické rozdíly. Na nejaridnějších ostrovech, kde jsou hojné stromové kaktusy, si endemické želvy vyvinuly dlouhé krky a přední končetiny a odlišně tvarované pancíře, které jim dovolují dosáhnout dostatečně vysoko, aby se mohly krmit těmito rostlinami. Na vlhčích ostrovech, kde se želvy nejčastěji krmí nižší vegetací, mají více zjednodušené tvary těla.
Tyto populace divergovaly z jednoho předka, který původně kolonizoval starší ostrovy v souostroví (dnes reprezentované ostrovy Española a San Cristóbal), pravděpodobně z Jižní Ameriky před cca 2 - 3 miliony let. Jeho potomci se rozšířili, a založily populace na mladších ostrovech. Vyskytly se nejméně dvě nezávislé kolonizace ostrovů Santa Cruz a Isabela. Každý z těchto ostrovů byl tak kolonizován vícekrát v různém čase a z různých zdrojových ostrovů. Ty ostrovy, které byly kolonizovány více než jednou mají alespoň dva pojmenované taxony, ačkoli taxonomové dosud neurčili, zda morfologické a molekulární rozdíly mezi různými populacemi galapážských želv nás opravňují považovat některé z nich za odlišné druhy nebo zda je lepší považovat je za poddruhy v rámci jediného druhu (Geochelone nigra nebo G. elephantopus).
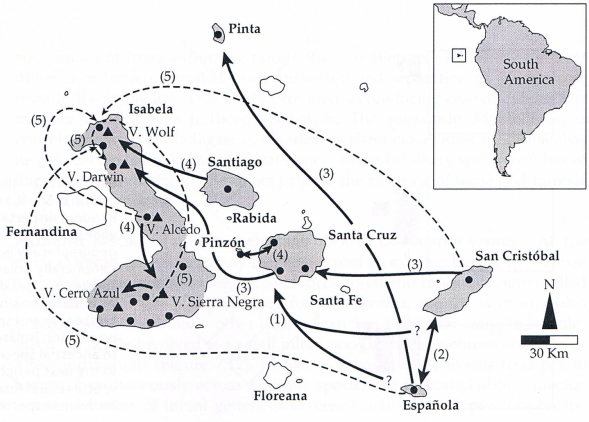
Obr. 33: Fylogeografická historie galapážských želv na větších ostrovech Galapážského souostroví. Pevninský předek z Jižní Ameriky nejprve kolonizoval starší ostrovy San Cristóbal a Española. Šipky ukazují směry pravděpodobné kolonizace; čísla označují přibližné pořadí kolonizací; plné šipky označují přirozené kolonizace a přerušované šipky označují kolonizace, které mohly být zapřičiněny lidmi. Poznamenejme, že se zde odehrály nejméně dvě nezávislé kolonizace ostrovů Santa Cruz a Isabela (Caccone et al. 2002, Lomolino et al. 2006).
Dalším příkladem jsou Darwinovy pěnkavky. I tady se věří, že všechny druhy jsou odvozeny z jediné populace předka, která kolonizovala souostroví před 2 - 3 miliony let, pravděpodobně z Karibských ostrovů. Avšak v případě pěnkavek, po divergování v izolaci na rozdílných ostrovech nebo na rozdílných přírodních stanovištích, některé populace úspěšně znovu pronikly do již osídlených areálů, takže několik druhů nyní koexistuje na jednom ostrově. Tyto druhy si vyvinuly nejen specifické chování při páření, které zabraňuje všeobecné mezidruhové hybridizaci, ale také divergovaly v morfologii (tvar zobáku podle specializace na určitý druh potravy), aby mohly využívat různé ekologické niky. Proces speciace pokračuje a jsou zde zdokumentovány případy hybridizace mezi formami, které dosud nejsou zcela reprodukčně izolované.

Obr. 34: Adaptivní radiace pěnkavek na Galapágách a rozmanitost jejich zobáků specializovaných na určitý typ potravy. Jeden z vlevo uvedených druhů pravděpodobně kolonizoval Galapážské souostroví z Karibských ostrovů. Následná allopatrická speciace zapřičiněná opakujícími se epizodami kolonizace a divergence vedla ke vzniku 5 rodů a 14 druhů (druh Cactospiza heliobates není uveden) (Lomolino et al. 2006).
Kontakt izolovaných populací a posílení speciace
Když se izoláty jednou zformují a diferencují, existuje několik možných závěrů: Za prvé mohou přijít opět do kontaktu s původním druhem. To se může stát buď zmizením geografické bariéry, nebo disperzí zpět přes ni. Pokud si populace znovu vytvořily kontakt, jsou tři možné výsledky:
1) Populace se nebudou křížit, protože si vyvinuly prezygotické izolační mechanismy nebo, pokud se kříží, nejsou schopné produkovat plodné potomstvo (postzygotický izolační mechanismus). V tom případě je reprodukční izolace úplná, proběhla speciace a druhy se mohou začít dělit o geografický prostor, ale jako dva oddělené druhy.
2) Obě populace se mohou značně křížit, až postupně geneticky splynou a jejich diferenciace se rozpadne.
3) Obě populace si nevyvinuly prezygotické izolační mechanismy a kříží se, ale hybridi jsou mimo úzkou hybridní zónu méně zdatní než potomci z páření v rámci populace. V tomto posledním případě selekce upřednostňuje ty jedince, kteří si vybírají partnery uvnitř vlastní populace a kteří budou dostatečně silní, aby vedly reprodukční izolaci a dokončení procesu speciace. Tento proces selekce prezygotického mechanismu, který podporuje páření uvnitř populace se označuje jako posílení speciace (Reinforcement, Dobzhansky 1937).
Posledním scénářem je, že izolované
populace nikdy nepřijdou do kontaktu. V tomto případě může
dlouho trvat, než se vyvine reprodukční izolace.
<< Předchozí | Nahoru | Následující >>
Zdroje:
BEGON, M., HARPER, J.L. & TOWNSEND, C.R. (1997): Ekologie: jedinci, populace a společenstva. 1. vyd. Vydavatelství Univerzity Palackého, Olomouc. 949 s. ISBN 8070676957
BROOKS, R.D. & McLENNAN (2002): The Nature of Diversity: An Evolutionary Voyage of Discovery. University of Chicago Press, Chicago.
BOSSUYT, F. & MILINKOVITCH, M.C. (2001): Amphibians as indicators of early Tertiary "out-of-India" dispersal of vertebrates. Science. 93-95.
CACCONE, A.G., GENTILE, G., GIBBS, J.P., FRITTS, T.H., SNELL, H.L., BETTS, J. & POWELL, J.R. (2002): Phylogeography and history of giant Galapagos tortoises. Evolution 56: 2052-2066.
COX, B.C. & MOORE, P. D. eds. (1999): Biogeography: an ecological and evolutionary approach. 6th edition. Oxford: Blackwell Science. ix, 298 s. ISBN 086542778X
KREBS, CH.J. (2001): Ecology: the experimental analysis of distribution and abundance. 5th ed. San Francisco, Calif.: Benjamin Cummings. 695 s. ISBN 0321042891
LOMOLINO, M. V., RIEDLE, B. R., & BROWN, J. H. eds. (2006): Biogeography. 3rd edition. Sunderland, Mass.: Sinauer Associates. xiii, 845. ISBN 0-87893-062-0
MACDONALD, G.M. (2003): Biogeography – Space, Time and Life. John Wiley & Sons, Inc. New York. ISBN 0-471-24193-8
