
3.2 Velikost a tvar areálu
Rozdíly ve velikostech areálů jednotlivých druhů jsou obrovské. Některé druhy jako například sokol stěhovavý (Falco peregrinus), sova pálená (Tyto alba) nebo orlovec říční (Pandion haliaetus) se vyskytují na všech kontinentech kromě Antarktidy. Na druhou stanu však většina druhů má areál menší než 5 000 000 km2 a jen málo druhů má areál větší než 10 000 000 km2. Bez ohledu na kontinent bylo zjištěno, že většina suchozemských živočiů má areály, menší než 1/5 až 1/4 velikosti kontinentu na kterém žijí. Obecně lze frekvenční rozložení velikostí areálu jednotlivých druhů popsat lognormální křivkou, která je unimodální a sešikmená vpravo. Toto typické rozložení četností skládající se z mnoha druhů s malými areály a jen z několika druhů s velkými areály se někdy také nazývá „hollow curve distribution“.

Obr. 45: Rozložení četností velikostí areálů ptáků Severní a centrální Ameriky (podle Brown, 1995).
Americký biogeograf James Brown pozoroval, že druhy s velkými areály se zpravidla objevují ve vysokých populačních hustotách v celém svém areálu, zatímco druhy s menšími areály se obvykle vyskytují v relativně nízkých hustotách. Předpokládá se tedy, že šířka ekologické niky druhu hraje klíčovou roli pro velikost areálu druhu a jeho populační hustotu. Brown naznačil, že běžné druhy můžou využít širokou nabídku prostředí a zdrojů, což jim umožní mít velký geografický areál. Podle metapopulační teorie jsou druhy, které si vytvořily velký areál s vysokou hustotou populace, silnější ve schopnosti šíření a v odolnosti proti vyhynutí. Vzhledem k tomu, že dílčí populace druhu jsou vzájemně prostorově oddělené, nebudou se katastrofické události nebo vypuknutí choroby v jedné z nich pravděpodobně týkat všech populací. Podle metapopulační teorie disponují tyto druhy také velkým množstvím migrujících jedinců, kteří rychle doplňují případné poklesy v okolních populacích.

Obr. 46: Vztah mezi velikostí areálů severoamerických druhů ptáků a hustotou jejich lokálních populací. Povšimněte si velmi malého počtu druhů s malými areály, ale vysokou populační hustotou (Brown 1995, Lomolino et al. 2006).
Porovnáním různých skupin organismů zjistíme, že druhy hmyzu a rostlin mají obecně menší průměrnou velikost areálu než obratlovci. Obojživelníci a sladkovodní ryby mají většinou menší areál, než savci a ptáci. Ptáci mívají větší areály než savci. Např. severoameričtí ptáci mají průměrnou velikost areálu přes 3 000 000 km2, zatímco severoameričtí savci mají průměrnou velikost areálu cca 1 500 000 km2. Ne všechny létavé organismy však mají rozsáhlý areál. Například průměrná velikost areálu netopýřích druhů v Severní a Střední Americe je přibližně stejná, jako v případě nelétajících savců.
Pokud se podíváme detailněji na ptáky a savce, zjistíme, že predátoři mají obecně větší areály než býložravci. V Africe mají velcí masožravci, jako lvi a hyeny, průměrnou velikost areálu 9 000 000 km2. Oproti tomu velcí býložravci, jako jsou sloni a zebry, mají velikost areálu kolem 4 000 000 km2. V Severní Americe je průměrná velikost areálu dravých sov okolo 6 000 000 km2, zatímco pro semenožravé vrabce je to kolem 3 500 000 km2. Relativně velké areály dravců pravděpodobně odráží jejich vyšší trofickou úroveň, neboť aby tito predátoři sehnali adekvátní potravu, musí být schopni lovit různé druhy kořisti v širokém rozpětí různých typů prostředí.
Zkoumání vztahu mezi velikostí těla zvířat a velikostí jejich areálu ukazuje, že velké areály mohou mít jak malé tak i velké druhy. Obvykle však druhy s většími těly vyžadují větší množství potravy, a proto potřebují větší plochu k obživě, než druhy s malými těly. Nicméně, existuje i velmi mnoho velkých druhů, které zaujímají jen velmi malé areály. Jestliže však velké druhy mají malý geografický areál, pak mají obvykle nízkou populační hustotu a jsou velmi náchylné k vyhynutí. Velký areál je proto pro přežití velkých druhů důležitý.

Obr. 47: Vztah mezi velikostí areálu severoamerických savců (A) a ptáků (B) a logaritmem velikosti těla. Povšimněte si velmi malého počtu velkých druhů s malými areály (Brown 1995, Lomolino et al. 2006).
Podstatný vliv na velikost areálu rostlin a živočichů má také evoluce. I menší krajinné prvky, jako je řeka nebo malé pohoří, mohou být překážkou pro šíření semen nebo pro pohyb malých nelétavých organismů. Přítomnost takových překážek omezuje tok genů mezi populacemi a podporuje tím opatrickou speciaci (vznik nových druhů vlivem izolace). Jejím důsledkem vznikají nové druhy, které mají zpočátku malé geografické areály. Určitý důkaz o roli krajinné rozmanitosti a fyzických překážek na utváření malých areálů poskytují biogeografické mapy. Severoameričtí a středoameričtí savci s areálem menším než 10 000 km2 jsou většinou malého vzrůstu a běžně se vyskytují v izolovaných horských oblastech, na ostrovech a poloostrovech. Členitý terén horských oblastí a vodní plochy obklopující ostrovy a poloostrovy tak vytvářejí překážky, které podporují alopatrickou speciaci, tedy vznik nových druhů s malými areály.
Areály rostlin a živočichů, kteří se vyskytují ve vysokých zeměpisných šířkách, jsou často větší, než areály druhů, které žijí v nízkých zeměpisných šířkách. Tento jev se nazývá Rapoportovo pravidlo a říká, že od rovníku k pólům se areály různých organismů zvětšují. Tento jev je pravděpodobně způsoben tím, že se zvětšující se zeměpisnou šířkou je lokální klima variabilnější, takže se druh musí adaptovat na různé klimatické podmínky a proto může osídlit velký geografický areál. Nicméně, důvody proč tomu tak skutečně je, se stále diskutují.

Obr. 48: Geografické rozložení velikosti areálů ptačích druhů. Pro každou buňku o velikosti 1° je velikost areálu vyjádřena jako medián velikostí areálů druhů vyskytujících se v dané buňce (Orme et al. 2006).
Gaston (2003) však ukazuje, že pro mnoho skupin organismů Rapoportovo pravidlo neplatí. U ptáků lze sice v holarktické oblasti vysledovat zvětšování areálů směrem na sever, ovšem v tropech jsou naopak areály spíše větší. Pro vysvětlení tohoto jevu Gaston opět využívá klima, ovšem jaksi obráceně: tropický pás zabírá největší část souše a má zároveň velmi stabilní teplotní podmínky, čímž je tropickému druhu poskytnut obrovský prostor s příznivým prostředím, kde může žít bez problémů ve velkém areálu.
Obecně lze říci, že tvary areálů rostlinných a živočišných druhů jsou spíše eliptické nebo obdélníkové, než kruhové či čtvercové. Protaženost areálu podél jedné osy může být řízena klimatem nebo reliéfem (pohořími apod.). Například severoamerické druhy s velkými areály mají často svůj areál protažený ve východo-západním směru. Naopak, druhy s malými areály vykazují spíše severojižní orientaci. Celkový geografický areál (reprodukční a zimní areál) kalypty růžovohlavé (Calypte anna) je omezen na úzký pás podél západního pobřeží Severní Ameriky. Naopak celý přirozený areál orla bělohlavého (Haliaeetus leucocephalus) se rozkládá od západního pobřeží Aljašky až k pobřeží Atlantického oceánu na východě Severní Ameriky. Jiná situace je však u evropských savců a ptáků, kde malé i velké areály jsou orientovány východo-západním směrem. Důvody těchto orientací je možné hledat ve fyzické geografii kontinentů. V Severní Americe i v Evropě mají hlavní vliv na uspořádání velkých areálů směrem východ-západ pásma klimatu a vegetace. Rozdíl v orientaci malých areálů na evropském a severoamerickém kontinentu je odrazem orientace pohoří (na severoamerickém kontinentu orientované severo-jižně zatímco v Evropě východo-západně), údolí, řek a pobřeží.
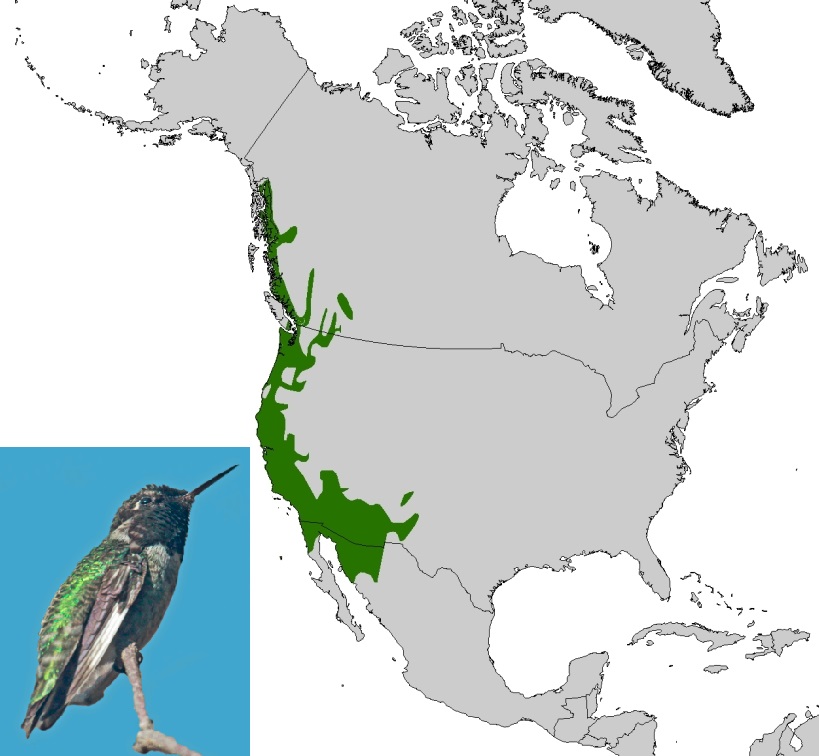
Obr. 49: Rozšíření kalypty růžovohlavé (Calypte anna; podle BirdLife International & Natureserve 2012; fotografie převzata z: http://commons.wikimedia.org/).

Obr. 50: Rozšíření orla bělohlavého (Haliaeetus leucocephalus; podle BirdLife International & Natureserve 2012; fotografie převzata z: http://commons.wikimedia.org/).
<< Předchozí | Nahoru | Následující >>
Zdroje:
BROWN, J. H., (1995). Macroecology. Chicago: University of Chicago Press.
BROWN, J. H. & MAURER, B. A. (1989): Macroecoogy: The division of food and space among species on continents. Science 243: 1145-1150.
BIRDLIFE INTERNATIONAL & NATURESERVE (2012): Bird species distribution maps of the world. BirdLife International, Cambridge, UK and NatureServe, Arlington, USA.
GASTON, K. J. (2003): The Structure and Dynamics of Geographic Ranges. Oxford: Oxford University Press.
LOMOLINO, M. V., RIEDLE, B. R., & BROWN, J. H. eds. (2006): Biogeography. 3rd edition. Sunderland, Mass.: Sinauer Associates. xiii, 845. ISBN 0-87893-062-0
MACDONALD, G. M. (2003): Biogeography – Space, Time and Life. John Wiley & Sons, Inc. New York. ISBN 0-471-24193-8
ORME, C. D. L. et al. (2006) Global Patterns of Geographic Range Size in Birds. PLoS Biology, 4, e208.
http://commons.wikimedia.org/wiki/Calypte_anna
http://commons.wikimedia.org/wiki/Haliaeetus_leucocephalus
